Лекция 20. Молекулярно-биологические основы возникновения жизни на Земле
|
Лекция 20. Молекулярно-биологические основы возникновения жизни на Земле |
|
Следующий раздел |
Список рисунков |
Эволюция пробиотов
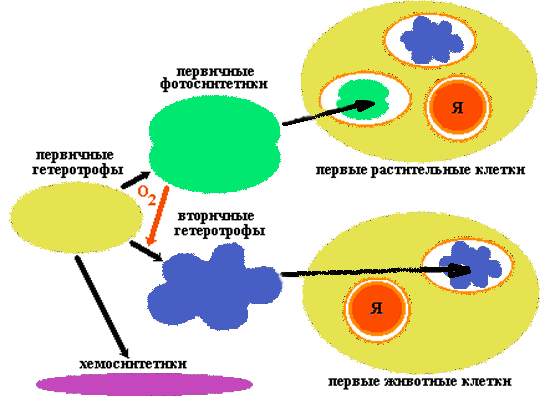 |
Пробионты были первичными гетеротрофами. Они получали энергию при расщеплении органических веществ абиогенного происхождения, в изобилии имевшихся в окружающей среде. Примером древнего способа обмена веществ, дошедшего до наших дней, является гликолиз - ферментативное бескислородное расщепление глюкозы.
По мере истощения запаса органического материала (а новый не образовывался из-за изменения условий на Земле) возникала жесткая конкурентная борьба за него, что ускорило процесс эволюции первичных гетеротрофов.
Исключительным событием стало возникновение бактериального фотосинтеза, освободившего клетки от зависимости от доступности органики абиогенного происхождения. Скорее всего, фотосинтез возник у анаэробных бактерий, способных к азотофиксации. Побочным продуктом фотосинтеза является кислород. Его накопление в атмосфере привело к коренному изменению хода эволюции. Появление озонового экрана защитило первичные организмы от смертельного УФ-облучения и положило конец абиогенному синтезу органики.
Первые аэробные бактерии появились благодаря приобретению аппарата окислительного фосфорилирования. Продукты брожения подвергались дальнейшему окислению до СО2 и Н2О. Аэробные (вторичные) гетеротрофы могли более эффективно, чем анаэробные (первичные) гетеротрофы, расщеплять органические вещества, образующиеся в результате фотосинтеза.
По-видимому, с ростом концентрации кислорода в атмосфере усложнялась жизнь первичных анаэробных гетеротрофов. Некоторые из них вымерли, другие нашли бескислородную среду. Примером могут служить дошедшие до наших дней метанобразующие бактерии или серные бактерии, живущие в горячих подземных источниках.
Некоторые первичные гетеротрофы пошли по пути, приведшему к образованию эукариотических клеток. Часть из них вступила в симбиоз с аэробными бактериями, способными к окислительному фосфорилированию. Поглотив вторичных гетеротрофов, первичные не расщепили их на молекулы, а сохранили в качестве энергетических станций, называемых сегодня митохондриями.
Такие симбионты дали начало царствам животных и грибов.
Другая часть первичных гетеротрофов "заключила союз" не только с аэробными гетеротрофами, но и с первичными фотосинтетиками, сохранив последних в качестве хлоропластов. Такие симбионты дали начало царству растений.
В пользу симбиотической теории образования эукариот говорят следующие факты:
- У митохондрий и хлоропластов две мембраны. Внутренняя - своя, наружняя образована клеткой-захватчиком.
- Генетический код митохондрий идеален. Универсальный генетический код имеет два существенных отличия, касающихся инициации и терминации синтеза белка.
Таким образом эукариоты отстранились от чужой генетической информации.
Кроме того, они линеаризовали свою ДНК. Митохондрии и хлоропласты имеют кольцевую ДНК, хотя не очень понятно, для чего им нужна кольцевая ДНК, и бактериальные рибосомы. Однако понятно, почему у них такая ДНК и такие рибосомы. Потому, что их предки были бактериями. Сегодня часть генов митохондриальных белков и белков хлоропластов, в том числе их РНК- и ДНК-полимераз, находятся в ядре. Вероятно, попали они туда с помощью мобильных элементов.
Все бактерии делятся на эубактерии (в том числе E.сoli) и археобактерии. Принципиальное отличие между ними в том, что гены археобактерий имеют экзон - интронное строение и сплайсинг. Эубактерии - результат эволюции ана- и аэробных гетеротрофов. Их эволюция шла в благоприятных условиях и они сменили больше поколений, избавившись от интронов. Археобактерии живут в экстремальных условиях: горячие, кислые, высокосолевые подземные воды. Эукариоты и археобактерии сохранили экзон - интронную структуру, что говорит о древнем происхождении экзонов и интронов.
|
Следующий раздел |
Список рисунков |