мЕЛГЙС 13. уИЕНЩ ТЕРМЙЛБГЙЙ дол in vivo
|
мЕЛГЙС 13. уИЕНЩ ТЕРМЙЛБГЙЙ дол in vivo |
| уРЙУПЛ ТЙУХОЛПЧ |
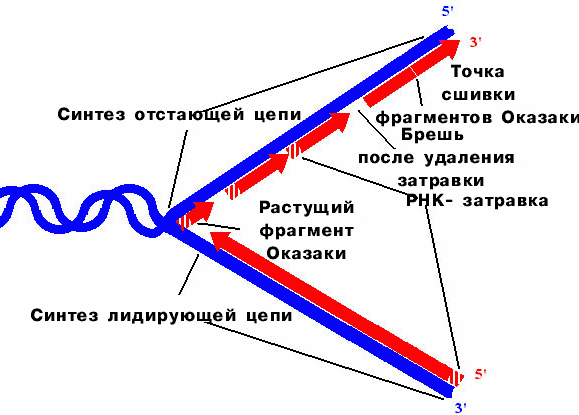
уИЕНБ РТЕТЩЧЙУФПК БОФЙРБТБММЕМШОПК ТЕРМЙЛБГЙЙ тЕКДЦЙ пЛБЪБЛЙ
1968 З.
йУИПДОЩЕ РПУЩМЛЙ: ЧУЕ ДБООЩЕ лПТОВЕТЗБ, РПМХЮЕООЩЕ Ч ЖЕТНЕОФБФЙЧОПК УЙУФЕНЕ in vitro, Й "ЛБТФЙОЛЙ" лЬТОУБ ЧЕТОЩ ( ОП ЛБТФЙОЛЙ ОЕРТБЧЙМШОП ЙОФЕТРТЕФЙТПЧБОЩ).
пЛБЪБЛЙ УРЕГЙБМШОП ТБЪТБВПФБМ ДЧБ ОПЧЩИ НЕФПДБ ЙУУМЕДПЧБОЙС.
1. нЕФПД ЙНРХМШУОПЗП НЕЮЕОЙС.
дП пЛБЪБЛЙ НЕФЛХ ДБЧБМЙ Ч ЛХМШФХТБМШОХА УТЕДХ Й ВЩУФТП ОБЮЙОБМЙ ПФНЩЧБФШ ЛМЕФЛЙ,
ОП НЙОЙНБМШОПЕ ЧТЕНС РПДБЮЙ НЕФЛЙ ВЩМП 5 НЙО. пЛБЪБЛЙ ЮЕТЕЪ ОХЦОЩК НПНЕОФ ЧТЕНЕОЙ
РПУМЕ ДПВБЧМЕОЙС НЕЮЕОПЗП ФЙНЙДЙОБ ДБЧБМ 1000-ЛТБФОЩК ЙЪВЩФПЛ ИПМПДОПЗП (ОЕНЕЮЕОПЗП)
ФЙНЙДЙОБ. фБЛЙН ПВТБЪПН, НЕФЛБ ЧЛМАЮБЕФУС ФПМШЛП Ч ФЕЮЕОЙЕ ПЮЕОШ ЛПТПФЛПЗП ЧТЕНЕОЙ.
пЛБЪБЛЙ УЮЙФБМ, ЮФП ЧТЕНС лЬТОУБ (5 НЙО.) ПЮЕОШ ЧЕМЙЛП ДМС РПМХЮЕОЙС ЙУФЙООПК ЛБТФЙОЩ РТПЙУИПДСЭЕЗП РТЙ ТЕРМЙЛБГЙЙ.
2. гЕОФТЙЖХЗЙТПЧБОЙЕ Ч ЭЕМПЮОПН ЗТБДЙЕОФЕ УБИБТПЪЩ.
уБИБТПЪБ ТБЪЧПДЙФУС ОБ ЭЕМПЮЙ. ч ЭЕМПЮОПК УТЕДЕ РТПЙУИПДЙФ ДЕОБФХТБГЙС дол.
ч ЬФПН УМХЮБЕ ЛПТПФЛЙЕ ЖТБЗНЕОФЩ дол, ЕУМЙ ПОЙ ЕУФШ, ПФДЕМСАФУС ПФ ДМЙООЩИ.
рПУМЕ ЬФПЗП ЙИ НПЦОП ЧЩСЧЙФШ РТЙ ГЕОФТЙЖХЗЙТПЧБОЙЙ Ч ЗТБДЙЕОФЕ РМПФОПУФЙ УБИБТПЪЩ,
ТБЪДЕМСАЭЕН НПМЕЛХМЩ РП НПМЕЛХМСТОПНХ ЧЕУХ.
пЛБЪБЛЙ РТЕДРПМПЦЙМ, ЮФП УЙОФЕЪ дол ЙДЕФ ЛПТПФЛЙНЙ ЖТБЗНЕОФБНЙ Й ЮФП ЛПТПФЛЙЕ ЖТБЗНЕОФЩ ДПМЦОЩ УЫЙЧБФШУС.
уЫЙЧБАФУС ПОЙ МЙЗБЪПК. дол-МЙЗБЪБ ВЩМБ ПВОБТХЦЕОБ ЛБЛ Х РТПЛБТЙПФ, ФБЛ Й Х ЬХЛБТЙПФ. х E. Уoli ВЩМЙ ОБКДЕОЩ НХФБОФЩ РП МЙЗБЪЕ.
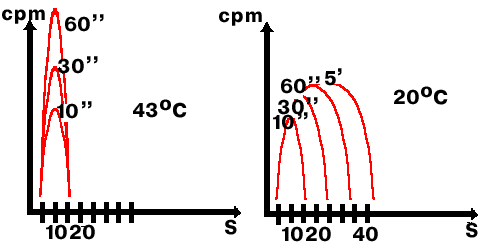
пЛБЪБЛЙ РТПЧЕМ ЬЛУРЕТЙНЕОФ ОБ ВБЛФЕТЙСИ, ЪБТБЦЕООЩИ ЖБЗПН ф4,
Х ЛПФПТПЗП ЕУФШ УЧПС ФЕТНПЮХЧУФЧЙФЕМШОБС МЙЗБЪБ, ЛПФПТБС ТБВПФБЕФ РТЙ 20![]() у
Й ОЕ ТБВПФБЕФ РТЙ 43
у
Й ОЕ ТБВПФБЕФ РТЙ 43![]() у.
еУМЙ Ч ЛМЕФЛХ РПРБДБЕФ ЖБЗПЧБС дол, ФП ЛМЕФЛБ РЕТЕЛМАЮБЕФУС ОБ ТЕРМЙЛБГЙА дол
ЖБЗБ.
у.
еУМЙ Ч ЛМЕФЛХ РПРБДБЕФ ЖБЗПЧБС дол, ФП ЛМЕФЛБ РЕТЕЛМАЮБЕФУС ОБ ТЕРМЙЛБГЙА дол
ЖБЗБ.
лМЕФЛЙ ЪБТБЦБМЙ ЖБЗПН ф4, ДБЧБМЙ ЙНРХМШУОХА НЕФЛХ о3-ФЙНЙДЙО
Й ЧЩТБЭЙЧБМЙ РТЙ ДЧХИ ФЕНРЕТБФХТБИ: 20![]() у
Й 43
у
Й 43![]() у.
рПФПН РТПЧПДЙМЙ ГЕОФТЙЖХЗЙТПЧБОЙЕ Ч ЭЕМПЮОПН ЗТБДЙЕОФЕ УБИБТПЪЩ.
у.
рПФПН РТПЧПДЙМЙ ГЕОФТЙЖХЗЙТПЧБОЙЕ Ч ЭЕМПЮОПН ЗТБДЙЕОФЕ УБИБТПЪЩ.
 |
|
мЙЗБЪБ E. coli ОХЦДБЕФУС Ч ЛПЖЕТНЕОФЕ обд, Б МЙЗБЪБ ЖБЗБ - Ч бфж. еУМЙ E.coli ОЕ ДБЧБФШ ОЙЛПФЙОПЧХА ЛЙУМПФХ (РТЕДЫЕУФЧЕООЙЛ обд), ВБЛФЕТЙБМШОБС дол ТЕРМЙГЙТХЕФУС, ОП ОЕ УЫЙЧБЕФУС.
рТЕТЩЧЙУФПУФШ ТЕРМЙЛБГЙЙ РПЛБЪБОБ ДМС ЧУЕИ ПВЯЕЛФПЧ, ЛТПНЕ ЖБЗПЧ, УПДЕТЦБЭЙИ ПДОПГЕРПЮЕЮОХА дол.
х ОЕЛПФПТЩИ ЖБЗПЧ Й ЧЙТХУПЧ РТЕТЩЧЙУФЩК УЙОФЕЪ ЙДЕФ РП ПВЕЙН ГЕРСН. х ВБЛФЕТЙК Й ЧЩУЫЙИ ПТЗБОЙЪНПЧ ПДОБ ГЕРШ ПВТБЪХЕФУС ОЕРТЕТЩЧОП, Б ДТХЗБС - РТЕТЩЧЙУФП (МЙДЙТХАЭБС Й ЪБРБЪДЩЧБАЭБС ГЕРЙ).
 |
тБЪНЕТ ЖТБЗНЕОФПЧ пЛБЪБЛЙ ЧЙДПУРЕГЙЖЙЮЕО Й УПУФБЧМСЕФ ДМС ЖБЗПЧ 1000-2000 ОХЛМ., E. Уoli - 1000 ОХЛМ., ДМС ЬХЛБТЙПФ - 200-400 ОХЛМ.
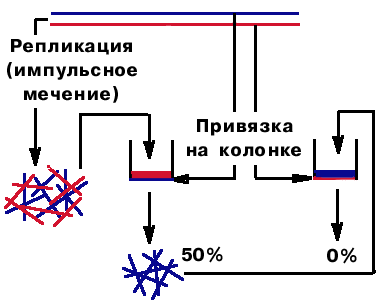 |
х ЖБЗБ ф7 ПВЕ ГЕРЙ дол ФПЦЕ ТЕРМЙГЙТХАФУС РТЕТЩЧЙУФП. пОЙ ЙНЕАФ ТБЪОХА РМПФОПУФШ (Ч ПДОПК ВПМШЫЕ РХТЙОПЧ). гЕРЙ ВЩМЙ ТБЪДЕМЕОЩ Ч ЗТБДЙЕОФЕ РМПФОПУФЙ CsCl Й РПНЕЭЕОЩ ОБ ДЧЕ ЛПМПОЛЙ. вЩМБ ПУХЭЕУФЧМЕОБ ЛПЧБМЕОФОБС РТЙЧСЪЛБ, РПЬФПНХ ПОЙ ОЕ НПЗМЙ УПКФЙ УП УНПМЩ. рПФПН РТПЧЕМЙ ТЕРМЙЛБГЙА Ч ХУМПЧЙСИ ЙНРХМШУОПЗП НЕЮЕОЙС. рТПРХУФЙМЙ ЧУЕ ЖТБЗНЕОФЩ ЮЕТЕЪ РЕТЧХА ЛПМПОЛХ. чЩИПД ВЩМ 50%. пУФБМШОЩЕ РТПЗОБМЙ ЮЕТЕЪ ЧФПТХА ЛПМПОЛХ. чЩИПДБ ОЕ ВЩМП. фБЛЙН ПВТБЪПН, ВЩМП РПЛБЪБОП, ЮФП ЖТБЗНЕОФЩ ЛПНРМЕНЕОФБТОЩ ПВЕЙН НБФТЙЮОЩН ГЕРСН. ъОБЮЙФ, ЖТБЗНЕОФЩ ПВТБЪХАФУС РП ПВЕЙН ГЕРСН |
| уРЙУПЛ ТЙУХОЛПЧ |