5
НЕРВНАЯ РЕГУЛЯЦИЯ ВИСЦЕРАЛЬНЫХ ФУНКЦИЙ. АВТОНОМНАЯ (ВЕГЕТАТИВНАЯ) НЕРВНАЯ
СИСТЕМА
ВЕХИ ИСТОРИИ
II в. — К. Гален (С. Galen, Др. Рим) дал название «симпатический» нервному стволу,
располагающемуся вдоль позвоночного столба, описал ход и распределение
блуждающего нерва.
1543 — А. Везалий (A. Vesalius, проф. в Падуе) дал подробное описание
симпатических узлов, ганглиев солнечного сплетения, полагая, что эти
образования осуществляют связь внутренних органов с мозгом.
1732 — Дж. Уинслоу (J. Winslow,
Великобритания) считал симпатические ганглии независимыми нервными центрами,
координирующими и согласующими функции различных органов.
1801 — Ф. Биша (F. Bichat, Франция) разделил жизненные процессы в организме на
животные и органические, полагая, что животная жизнь зависит от спинного мозга
и является соматической, в то время как органическая поддерживается
симпатической ганглионарной системой. Он также считал, что симпатические узлы
наделены самостоятельной деятельностью.
1807 — И. Рейл (I. Real, Великобритания) ввел понятие «вегетативная нервная
система».
1833, 1836 — В. Эленберг и Г. Валентин (W. Elenberg, G. Valentin, Германия)
представили первое микроскопическое описание ганглионарной нервной клетки.
1836 — И. Браше (I. Brachet, Франция) издал монографию о
функциях ганглионарной нервной системы, в которой утверждал, что эта система
регулирует деятельность внутренних органов и описал ее влияния на сердце,
легкие, тонкую кишку, органы выделения, органы чувств.
1840 — А. М. Филомафитский (Россия)—различал в симпатическом
нерве такие свойства, как чувствительность, двигательную и органическую силу.
Наличие двигательной силы он подтверждал раздражением сердечного нерва
электрическим током, в результате чего наблюдались сильные движения сердца.
Кроме того, ему первому удалось показать симпатические влияния на химические
процессы в организме.
1845 — братья Э. Г. и Э. Веберы (Er. H. и Ed. Weber, Германия)
обнаружили тормозящее влияние блуждающего нерва на деятельность сердца. Это
было первое исследование о торможении (угнетении) физиологических процессов.
1852 — К. Бернар (С. Bernard, Франция) описал сосудодвигательную функцию симпатических
нервов. В дальнейшем его концепция о значении постоянства внутренней среды
организма послужила основой для формирования учения о гомеостазе.
1857, 1864 — Г. Мейсснер и Л. Ауэрбах (G. Meissner, L. Auerbach, Германия)
представили описание гистологического строения подслизистого и межмышечного
сплетения стенки кишки.
1858 — М. Шифф (М. Schiff, Германия)
описал специальные нервы регуляции питания что создавало почву для последующего
развития представлений о трофической функции симпатической нервной системы
1866 — И. Ф. Цион (Россия) и К. Людвиг (К. Ludwig, Германия) открыли у кролика нерв депрессор,
раздражение центрального конца которого вызывает рефлекторное падение кровяного
давления и расширение кровеносных сосудов.
1886 — И. П. Павлов (Россия) высказал предположение, что
блуждающий и симпатический нервы оказывают влияние не только на сокращение
сердца, но и изменяют питание (трофику) сердечной мышцы.
1886— У. Гаскелл (W. Gaskell, Великобритания) ввел термин
«висцеральная нервная система», понимая под этим иннервацию внутренних органов,
сосудов и желез.
1377 — А. И. Бабухин (Россия) описал аксон—рефлекс — рефлекс,
осуществляющийся по разветвлениям аксона без участия тела нервной клетки;
возбуждение при аксон—рефлексе может распространяться по разветвлениям
эфферентных симпатических, а также спинальных чувствительных волокон.
1893, 1898,
1922 — Дж. Ленгли (J. Langley, Великобритания) в 1893 г.
ввел в термины «пре— и постганглионарные волокна». В 1898 г. впервые довольно
точно для своего времени описал общий план строения вегетативной нервной
системы, определил места выхода вегетативных волокон из центральной нервной
системы, установил их анатомические и функциональные особенности, подразделил
вегетативную нервную систему на симпатический и парасимпатические отделы,
выделил в самостоятельный отдел энтеральную систему (enteric system) —
мейсснерово и ауэрбахово сплетения. Назвал вегетативную нервную систему
автономной. В 1922 г. выпустил книгу «Автономная нервная система».
1921—1936 — О. Леви (О. Loewi, Австрия) установил, что при раздражении симпатического или
блуждающего нервов изолированного сердца лягушки в перфузате появляются
вещества, оказывающие соответственно симпатический или парасимпатический
эффекты. В 1936 г. вместе с Г. Дейлом (Н. Dale, Великобритания) удостоен Нобелевской премии «за открытия,
связанные с химической передачей нервных импульсов».
1923 — А. Г. Гинецинский (СССР) установил, что под влиянием
импульсов, идущих по симпатическим нервам, улучшается функция утомленной мышцы
в результате длительного раздражения двигательного нерва лягушки; эффект
воспроизведен и на мышцах млекопитающих; синоним — феномен Орбели—Гинецинского.
1932 — Л. А. Орбели (СССР) обосновал и развил учение о
адаптационно—трофической функции симпатической нервной системы,
распространяющееся на скелетные мышцы, рецепторы, всю ЦНС, включая кору больших
полушарий мозга.
1933
— А. В. Кибяков (СССР) установил роль адреналина в синаптической передаче.
1943, 1960, 1973 — В. Н.
Черниговский (СССР) создал учение об интероцепции; в 1943 г. издал книгу
«Афферентные системы внутренних органов», в 1960 г. — книгу «Интероцепторы», в
1973 г. описал кортикальное и субкортикальное представительство висцеральных
систем.
1980 — А. Д. Ноздрачев
экспериментально обосновал существование третьей части автономной нервной
системы — метасимпатической нервной системы (МНС).
Cогласно
представлениям французского врача и физиолога Ф. Биша (начало XIX столетия)
функции организма позвоночных разделяли на две группы: соматической
(анимальной, животной) и вегетативной (растительной) жизни. Первая включала
двигательные реакции скелетной мускулатуры и восприятие внешних раздражении, ко
второй причислялись все остальные функции тела — питание, рост, размножение и
т. д. В соответствии с этим был установлен сохранившийся до сих пор и
физиологический принцип разделения нервной системы на соматическую и вегетативную (от лат. vegetativus — растительный).
Позже
знаменитый французский физиолог К. Бернар постулировал новый признак
вегетативной нервной системы, превратив ее в систему непроизвольной
иннервации. Признак непроизвольности во многих отношениях оказался
плодотворным. Он позволил английскому физиологу У. Гаскеллу обратить внимание
на наличие в организме двух типов мышечной ткани, подчиненных соответственно
«произвольной» и «непроизвольной» иннервации.
В пределах
непроизвольной оказалась иннервация мышц сосудов, кожных образований,
внутренних органов. Признак непроизвольности оказался не единственным при
отнесении нервного образования к тому или иному отделу. Гаскелл показал также
существование своеобразной химической чувствительности мышц,
иннервируемых непроизвольной нервной системой, часть из которых реагирует
сокращением на применение адреналина. Это позволило ему разделить
непроизвольную нервную систему на собственно симпатическую (адреналовую)
и нервную систему внутренностей, выделив при этом краниальный, тораколюмбальный
и сакральный отделы.
Вскоре соотечественник
Гаскелла Дж. Ленгли установил различие в конструкции произвольной и
непроизвольной иннервации. Он показал, что произвольная соматическая иннервация
осуществляется однонейронным путем — тело нервной клетки лежит в ЦНС, а
ее отросток, располагаясь на периферии, достигает исполнительного органа
(скелетной мышцы). В то же время путь непроизвольной вегетативной иннервации
представлен двумя нейронами первый из которых находится в ЦНС, второй —
в периферическом ганглии.
Эту непроизвольную часть нервной
системы Ленгли назвал автономной нервной системой, подчеркнув тем самым
гораздо большую степень ее независимости от ЦНС. Он считал эту систему сугубо
эфферентной. В автономной нервной системе Ленгли выделил симпатическую
нервную систему (тораколюмбальный отдел) и обозначил остальные образования
(краниальный и сакральный отделы) термином парасимпатическая. При этом
нервные сплетения стенки кишки он отдельно назвал энтеральной системой.
Особую роль автономной
нервной системы в организме обосновал Л. А. Орбели. Данные его школы показали,
что симпатическая иннервация оказывает воздействие на функциональное состояние
всех без исключения органов и тканей, в том числе и отделов ЦНС. Таким образом
был сформулирован основной принцип ее функционирования в организме — адаптационно—трофический
характер оказываемого влияния.
Согласно международной
анатомической номенклатуре, сейчас термин автономная заменил все ранее
существовавшие —растительная, висцеральная, вегетативная и т. д. В
понятие автономная нервная система включается комплекс центральных и
периферических клеточных структур, регулирующих
функциональный
уровень внутренней жизни организма, необходимый для адекватных реакций всех
систем.
Анатомически автономная
нервная система высших позвоночных представлена ядерными образованиями,
лежащими в спинном и головном мозгу нервными ганглиями и нервными волокнами,
локализованными либо вблизи исполнительных органов, либо непосредственно в их
стенках или толще.
5.1. ДУГА АВТОНОМНОГО РЕФЛЕКСА
Как и
соматическая рефлекторная дуга, дуга автономного рефлекса состоит из
трех звеньев: чувствительного (сенсорного, афферентного), ассоциативного
вставочного) и эффекторного. Например, в автономной рефлекторной дуге спинального
уровня (рис. 5.1) чувствительное звено образовано клетками спинномозговых или
периферических ганглиев. Оно может быть общим для автономной и соматической
рефлекторных дуг.
Периферические отростки чувствительных клеток
разветвляются во внутренних органах, коже, стенках сосудов и т. д., центральные
же синаптически контактируют со вставочными нейронами тех или других сегментов.
Втopoе звено этой же дуги может быть упрощенно представлено в виде скопления
нейронов в боковых рогах спинного мозга. Их отростки покидают спинной мозг в
составе вентральных корешков, вступают в соматические нервные стволы и отсюда в
виде белых соединительных ветвей направляются к узлам симпатического ствола.
Здесь происходит переключение части из них на эффекторные
|
|
|
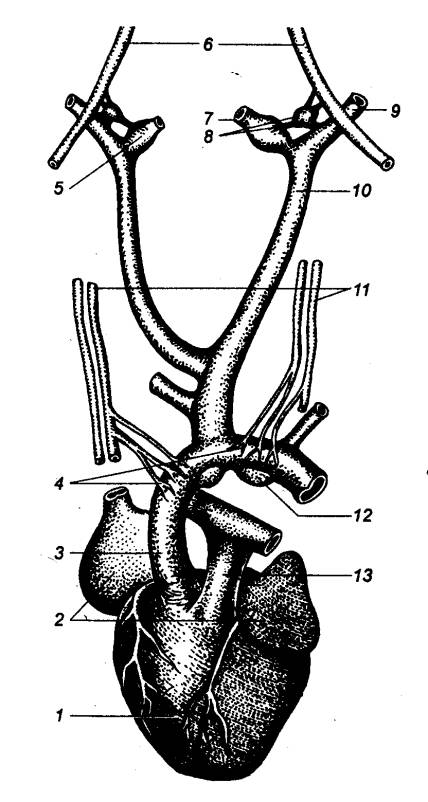
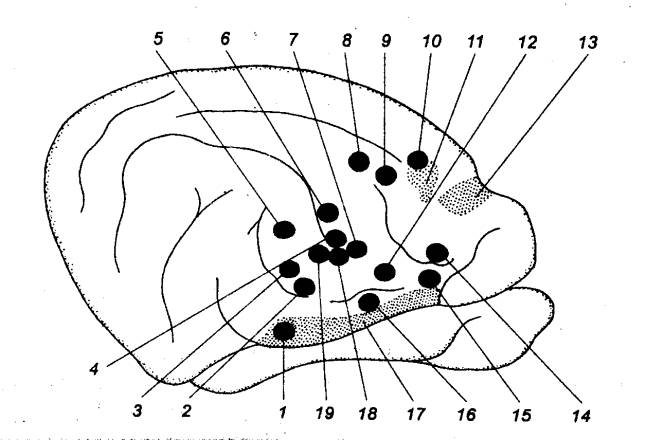
Рис. 5.1 Принципиальная
схема дуги автономного (справа) и соматического (слева) рефлексов 1 — спинной
мозг, 2 — чувствительный
нейрон, 3 — спинальный ганглий (спинномозговой узел), 4 —
дорсальный корешок, 5 — вставочный (преганглионарный) нейрон дуги автономного
рефлекса, 6 — вентральный
корешок, 7 — преганглионарное (предузловое) нервное волокно, 8 — симпатический паравертебральный
ганглий (узел симпатического ствола), 9
— белая соединительная ветвь, 10
— двигательный (постганглионарный) нейрон дуги автономного рефлекса, 11 — соматический нерв, 12 — постганглионарное
(послеузловое) нервное волокно, 13 —
серая соединительная ветвь, 14 —
висцеральная ветвь, 15 — симпатический
превертебральный ганглий (узел брыжеечного сплетения), 16 — кишка, 17 — эффекторный нейрон функционального модуля
метасимпатической нервной системы, 18
— функциональный модуль метасимпатической нервной системы, 19 — интернейрон функционального
модуля, 20 — чувствительный
нейрон функционального модуля, 21 —
двигательное соматическое волокно, 22
— двигательный нейрон соматической рефлекторной дуги, 23 — вставочный нейрон соматической
рефлекторной дуги. |
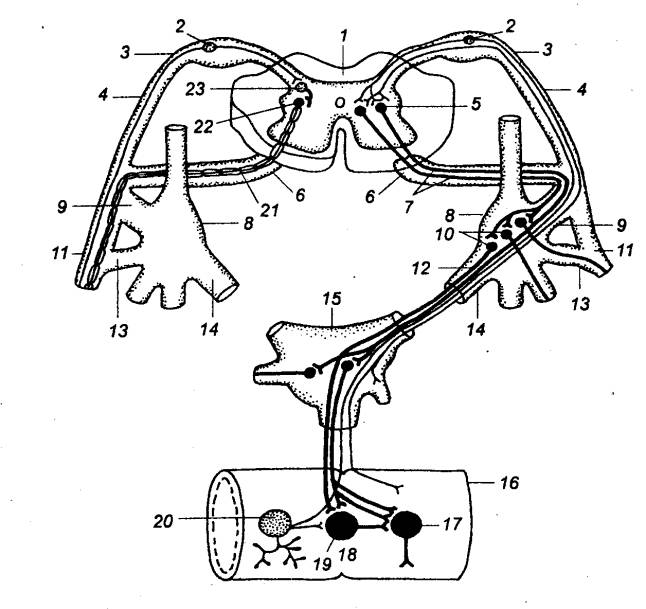
клетки. Третье звено — нервная клетка, мигрировавшая из
спинного мозга в один из периферических узлов.
Узлы могут
располагаться либо около позвоночника (паравертебральные), либо в
нервных сплетениях вблизи внутренних органов (превертебральные), либо,
наконец, в стенках внутренних органов (интрамуральные).
Дуги местных
рефлексов ганглионарного уровня могут замыкаться в превертебральных и
интрамуральных ганглиях, так что в этом случае все составляющие рефлекторную
дугу звенья находятся на периферии вблизи или непосредственно в стенке
исполнительного органа. При этом чувствительное и эфферентное волокна могут
проходить в составе одного и того же нервного ствола.
5.1.1.
Подразделение автономной нервной системы
Автономную нервную систему
подразделяют на симпатическую, парасимпатическую и метасимпатическую
части, или отделы (рис. 5.2). Это деление имеет определенные функциональные и
структурные основания. Они относятся ко всем трем основным звеньям рефлекторной
дуги. Согласно А. А. Заварзину, в процессе развития при появлении билатеральной
симметрии и дифференцировки головного и хвостового концов началось разделение
нервной системы на соматическую и автономную. Из этой системы выделилась часть
основного трехчленного аппарата, сохранившая свою связь с соматической
системой. В связи с более однообразной и менее сложной организацией
висцеральных органов, чем органы движения, появились особые свойства,
развившиеся из общих свойств единой нервной системы. Таким путем возникла
симпатическая, парасимпатическая и метасимпатическая нервная система.
В свою очередь, развитие частей
автономной нервной системы шло, вероятно, параллельно, чем объясняется наличие
у каждой из них единого принципа — параллельно поставленного аппарата. В
процессе эволюции в этом аппарате выработались особые свойства, характерные для
каждой из частей, но развившиеся из единой нервной системы. Так, в
метасимпатической части обособился сенсорный аппарат, возникли собственный
водитель ритма, интернейронная обрабатывающая сеть и эффекторный нейрон с
собственным медиаторным
|
|
|
1— крестцовый отдел спинного мозга (S), 2 — грудо—поясничный отдел спинного мозга (T—L) 3 — шейный отдел спинного мозга (С), 4— продолговатый
мозг, 5— средний мозг, 6— симпатический ствол (паравертебральные ганглии), 7—
парасимпатические узлы головы, 8— постганглионарные волокна, 9— (чревное)
солнечное сплетение и его узлы, 10— каудальное брыжеечное сплетение и его
узлы, преганглионарные нервные волокна, 12— тазовый нерв, 13— слюнные железы,
14— внутренние мышцы глаза, 15— сосуды головного мозга и его оболочек, 16—
трахея, бронхи, легкое, 17—функциональные модули метасимпатической нервной
системы, 18— желудок, 19— сердце, 20— скелетная мышца, 21— двенадцатиперстная
кишка, 22— поджелудочная железа, 23— сосуды внутренних органов, 24— печень,
25— надпочечник, 26— толстая кишка, 27— почка, 28— мочевой пузырь, 29—
семенники, 30— матка, 31— кожа |
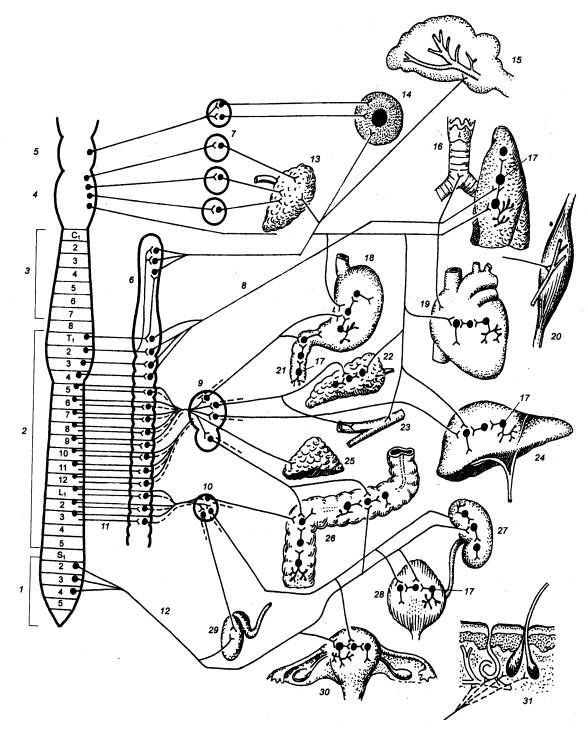
обеспечением.
Иными словами, возникли низшие рефлекторные центры, расположенные
непосредственно в стенках исполнительных органов. У симпатического и
парасимпатического отделов появились дуги с участием спинного мозга и
образованием собственных центров — ядер спинного и головного мозга, а также
высшего центра — гипоталамической области.
Эмбриональным источником клеток автономной нервной системы у млекопитающих
служит ганглиозная пластинка, которая подразделяется на сомиты, дающие
впоследствии симпатическую и парасимпатическую нервную систему. Их
периферическая часть, а также метасимпатическая нервная система образуются в
результате дальнейшей миграции нейробластов в стенки внутренних органов.
5.1.2. Анатомические структуры
Симпатическая нервная система. Симпатическая нервная система по
строению делится на центральную часть, расположенную в спинном мозгу, и
периферическую, включающую многочисленные ветви и узлы.
Центральная часть представлена симпатическим ядром бокового рога серого
вещества спинного мозга. Оно тянется от первых грудных до поясничных сегментов.
Аксоны составляющих ядро клеток через межпозвоночные отверстия выходят из
спинного мозга в составе вентральных корешков (см. рис. 5.1) и в виде белых
соединительных ветвей вступают в узлы симпатического ствола. Ядро состоит из
сравнительно мелких мультиполярных клеток, дендриты которых ветвятся здесь же,
а тонкие и часто миелинизированные аксоны, называемые преганглионарными
(предузловыми) волокнами, направляются в вентральные корешки и
оканчиваются либо в паравертебральных (околопозвоночных), либо в
превертебральных (предпозвоночных) ганглиях. Скорость проведения возбуждения в
этих аксонах колеблется от 1 до 20 м/с.
Периферическая часть образована чувствительными и эфферентными нейронами
симпатических ганглиев. В их анатомическом строении у высших позвоночных
животных и человека принципиальных отличий не имеется.
Паравертебральные ганглии располагаются по обе стороны позвоночника, от
основания черепа до крестца, в виде цепочек, называемых правым и левым
пограничными симпатическими стволами. Узлы между собой соединены
комиссурами, а со спинномозговыми нервами — белыми и серыми соединительными
ветвями (рис. 5.2, 5.3).
По белой ветви в узел входит преганглионарное волокно,
которое может переключаться здесь на эффекторный ганглионарный нейрон, тогда
часть постганглионарных волокон по серой соединительной ветви вновь
возвращается в спинномозговой нерв и далее следует в его составе без перерыва к
эффекторному органу, например, к скелетной мышце или суставу. Другие волокна,
собравшись в отдельные веточки, называемые внутренностными (чревными)
нервами, направляются к органам грудной, брюшной и тазовой полости или к
превертебральным ганглиям и через них далее также к исполнительным органам.
Постганглионарные волокна в большинстве лишены миелиновой оболочки, поэтому
имеют сероватую окраску. В ганглиях симпатического ствола прерывается только
часть преганглионарных волокон, остальные проходят их транзитом и синаптически
переключаются на эффекторный нейрон в превертебральных ганглиях.
Превертебральные ганглии расположены на значительном удалении от спинного
мозга и вдали от иннервируемых органов, поэтому от них идут довольно длинные
постганглионарные аксоны, совокупность которых в каждом случае носит название
специального нерва. Исключение составляют небольшие узлы,
|
|
|
Рис. 5.3 Симпатический и метасимпатический
отделы автономной нервной системы, (схема) Преганглионарные симпатические волокна обозначены сплошными линиями,
постганглионарные — пунктирными. Функциональные модули метасимпатической
нервной системы (МНС) показаны встроенными непосредственно в стенки полых
внутренних органов; местом локализации МНС являются интрамуральные ганглии. |
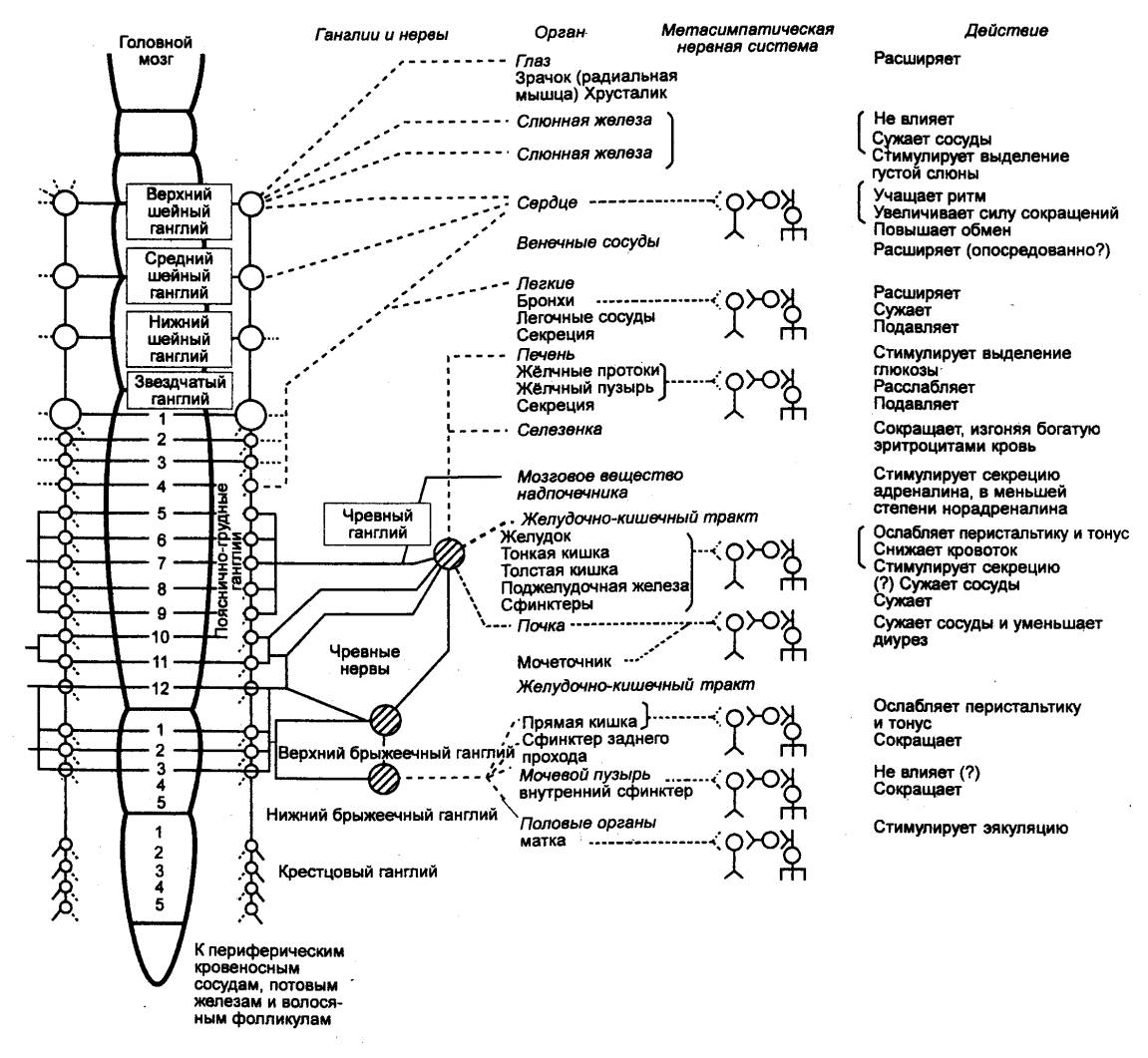
находящиеся вместе с половыми органами и отдающие им
короткие постганглионарные аксоны.
К числу наиболее крупных
превертебральных ганглиев относятся чревный, краниальный и каудальный
брыжеечные узлы. Два первых вместе с отходящими ветвями образуют самое крупное
из автономных сплетений организма — чревное сплетение, традиционно
называемое солнечным сплетением (рис. 5.2). От клеток этих узлов начинаются
постганглионарные симпатические волокна, иннервирующие почти все органы брюшной
полости. Они образуют многочисленные нервные стволы, которые отходят и
распределяются радиально. Стволы направляются к органам брюшной полости и в
свою очередь образуют ряд сплетений: печеночное, селезеночное, желудочное и др.
К эффекторам, снабжаемым
постганглионарными симпатическими волокнами, относятся гладкие мышцы всех
органов — сосудов, зрачка, волосяных луковиц, легких, органов пищеварения,
выделения, потовые, слюнные, пищеварительные железы, а также клетки печени и
жировой клетчатки.
Помимо эфферентных волокон
симпатическая нервная система имеет собственные чувствительные пути. По
локализации клеточных тел, ходу и длине отростков они могут быть разделены на
две группы. Первая группа
|
|
|
Рис. 5.4 Парасимпатический и
метасимпатический отделы автономной нервной системы, (схема) Холинергические волокна, идущие
из краниального и крестцового отделов ЦНС, направляются к ганглиям, органам и
к функциональным модулям метасимпатической нервной системы, локализующейся в
интрамуральных ганглиях стенок полых внутренних органов. |
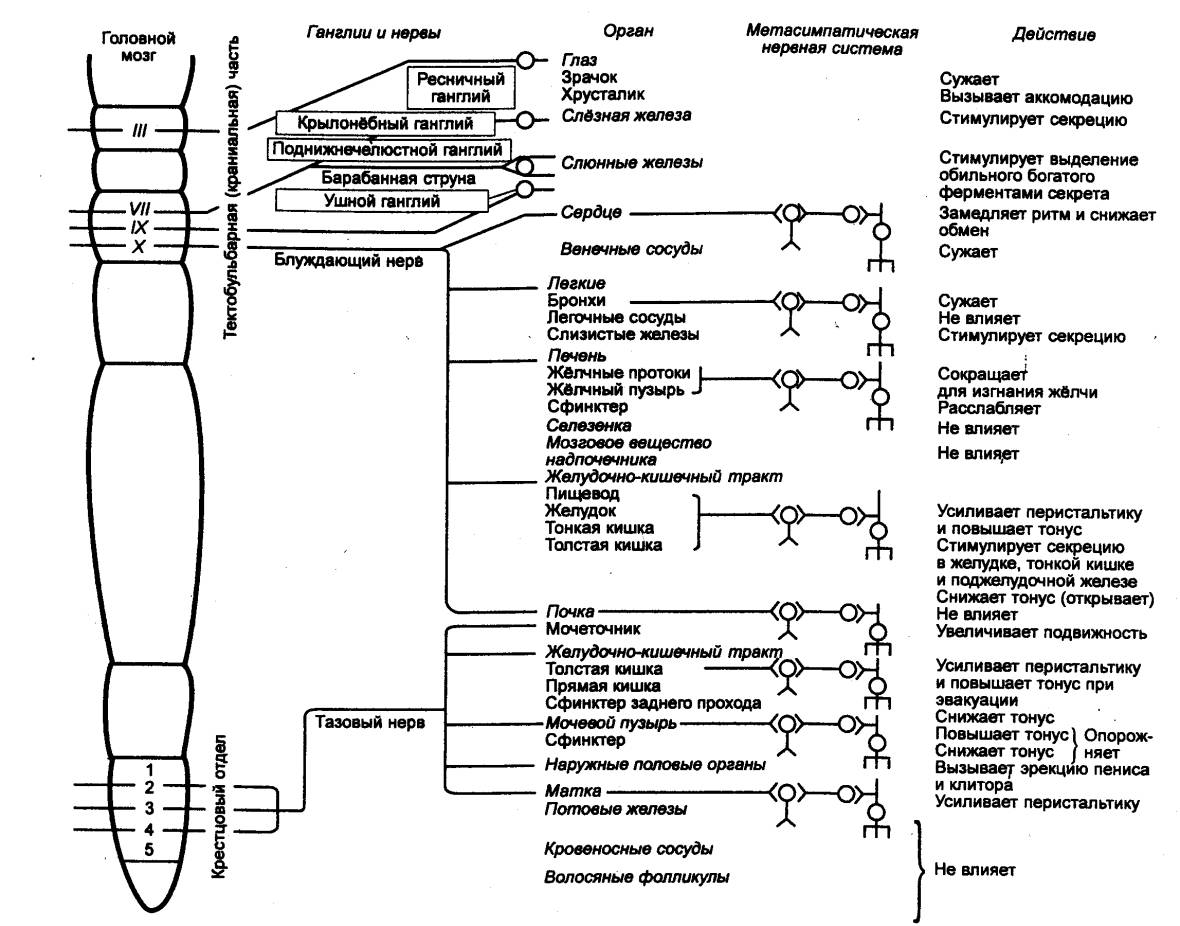
периферических афферентных нейронов включает клетки, тела
которых локализуются в превертебральных симпатических ганглиях. Один из длинных
отростков направляется на периферию, второй — в сторону спинного мозга, куда он
вступает в составе дорсальных корешков (см. рис. 5.1). Вторая группа
характеризуется тем» что длинный отросток этих чувствительных клеток идет к
рабочему органу, короткие же распределяются в самом ганглии, синаптически
контактируют с вставочными нейронами и через них с эффекторными нейронами, образуя
здесь местную рефлекторную дугу.
Парасимпатическая нервная система. Общая
организация парасимпатической нервной системы в какой—то мере подобна
симпатической. В ней также выделяются центральные и периферические образования,
передача возбуждения к исполнительному органу осуществляется в основном по двухнейронному
пути, преганглионарный нейрон располагается в сером веществе мозга,
постганглионарный вынесен далеко на периферию. Однако парасимпатическая нервная
система отличается от симпатической рядом особенностей. Во—первых, ее
центральные структуры расположены в трех различных далеко отстоящих участках
мозга, отделенных не только друг от друга, но и от симпатического центра; во—вторых,
парасимпатические волокна иннервируют, как правило, только определенные зоны тела,
которые также снабжаются симпатической, а иные и метасимпатической иннервацией.
Центральные структуры парасимпатической нервной системы
расположены в среднем, продолговатом мозгу и в крестцовом отделе спинного мозга
(рис. 5.2, 5.4).
Среднемозговая часть представлена парасимпатическим ядром
глазодвигательного нерва (ядром Якубовича, или Вестфаля—Эдингера), лежащим
вблизи краниальных (верхних) холмиков на дне водопровода среднего мозга
(сильвиевого). Преганглионарные волокна к глазным мышцам, железам и другим
образованиям головы покидают ствол мозга в составе трех пар черепных нервов:
III (глазодвигательного), VII (лицевого), IX (языкоглоточного) — и
заканчиваются на эффекторных нейронах ресничного, ушного, крылонёбного,
поднижнечелюстного (подъязычного) узлов. Отсюда постганглионарные волокна идут
к иннервируемым органам.
|
|
|
Рис. 5.5 Иннервация каротидной и аортальной
сосудистых зон 1 —
желудочки, 2 — предсердия, 3
— восходящая аорта, 4 —
барорецепторные области дуги аорты, 5 — сонный (каротидный) синус, 6 — языкоглоточные нервы, 7 —
внутренняя сонная артерия, 8 —
каротидные нервы, 9 —
наружная сонная артерия, 10 —
общая сонная артерия, 11 —
главные стволы блуждающих нервов, 12
— парааортальные тельца, 13 —
легочная артерия. |
Клеточные тела преганглионарных нейронов продолговатого
мозга посылают свои волокна к органам шеи, грудной и брюшной полости в составе
Х пары (блуждающего нерва). Крестцовый отдел представлен центрами,
располагающимися в боговых рогах трех крестцовых сегментов спинного мозга.
Отсюда в составе тазового внутренностного нерва парасимпатические волокна
направляются к органам таза.
Постганглионарные парасимпатические волокна снабжают глазные
мышцы, слёзные и слюнные железы, мускулатуру и железы пищеварительного тракта,
трахею, гортань, легкие, предсердия, выделительные и половые органы. В отличие
от симпатических постганглионарных волокон они не иннервируют гладкие мышцы
кровеносных сосудов, за исключением половых органов и, вероятно, артерий мозга.
Главным коллектором чувствительных путей парасимпатической
нервной системы является блуждающий нерв. При общем количестве волокон
его шейного отдела, достигающем, например, у кошки 30 тыс., 80—90% составляют афферентные
волокна. Примерно 20% из этого числа приходится на долю миелинизированных
волокон, остальные — тонкие безмякотные. Эти волокна передают информацию от
пищеварительного тракта, органов грудной и брюшной полости. Образуемые этими
волокнами рецепторы реагируют на механические, термические, болевые
воздействия, воспринимают изменения рН и электролитного состава.
Исключительно важна физиологическая роль чувствительной
ветви блуждающего нерва — депрессорного нерва (нерва, понижающего
кровяное давление). Он является мощным проводником, сигнализирующим о
функциональном состоянии сердца. Клеточные тела афферентных путей блуждающего
нерва лежат в основном в яремном узле, а их волокна вступают в продолговатый
мозг на уровне олив.
В синусной ветви (ветви сонного синуса), являющемся
ветвью IX пары (языкоглоточного нерва), проходит около 300 толстых волокон,
связанных с большим числом рецепторных приборов разной модальности. В этом
воспринимающем комплексе особое значение принадлежит сонному гломусу
(каротидному клубочку), лежащему между внутренней и наружной сонными артериями
у места деления общей сонной артерии (рис. 5.5).
Метасимпатическая нервная система. Многие внутренние органы после
перерезки симпатических и парасимпатических путей или даже после извлечения из
организма продолжают осуществлять присущие им функции без особых видимых
изменений. Сохраняет координированную перистальтическую и всасывательную
функции кишка, сокращается перфузируемое сердце. Сегменты или полоски матки,
мочеточника, жёлчного пузыря продолжают сокращаться с частотой и амплитудой,
характерной для каждого органа. Эта функциональная автономия объясняется
наличием в стенках этих органов ганглиозной системы, обладающей собственным
автоматизмом и имеющей необходимые для автономной рефлекторной и интегративной
деятельности звенья — чувствительное, вставочное, двигательное и медиаторы.
Следовательно, большинство полых висцеральных органов наряду
с существованием экстраорганной симпатической и парасимпатической нервной системы
имеет собственный базовый механизм нервной регуляции. Управление работой в этом
случае обеспечивается рефлекторными дугами, замыкающимися в пределах стенки
самих органов (рис. 5.6).
Наличие общих черт в структурной и функциональной
организации, данные онто— и филогенеза, результаты наблюдений за конечными
эффектами, возникающими при стимуляции нервных волокон, результаты прямой
регистрации нейрональной активности явились основанием для выделения в составе
автономной нервной системы кроме симпатической и парасимпатической еще и
третьей части — метасимпатической.
Ранее к третьему отделу автономной нервной системы,
называемому энтеральным, относили лишь рефлекторные дуги, замыкающиеся в
подслизистом и межмышечном сплетениях кишки. Понятие метасимпатической нервной
системы значительно шире, оно охватывает весь комплекс полых висцеральных
органов, в том числе и пищеварительный тракт. Местом локализации
метасимпатической нервной системы являются интрамуральные ганглии, залегающие в
толще стенок этих органов.
Метасимпатическая нервная система обладает многими
признаками, отличающими ее от других частей автономной нервной системы.
|
|
|
Рис.
5.6 Функциональный модуль
метасимпатической нервной системы 1 —
чувствительный нейрон, 2 —
интернейрон, 3 — эфферентный нейрон, 4
— постганглионарный симпатический нейрон и его волокно, 5 —
преганглионарный симпатический нейрон и его волокно, 6 — преганглионарный парасимпатический нейрон и его волокно. |
1. Она иннервирует только внутренние
органы, наделенные собственной моторной активностью; в сфере ее иннервации
находятся гладкая мышца, всасывающий и секретирующий эпителий, локальный
кровоток, местные эндокринные элементы, иммунные структуры.
2. Она получает синаптические
входы от симпатической и парасимпатической систем и не имеет прямых
синаптических контактов с эфферентной частью соматической рефлекторной дуги.
3. Наряду с общим висцеральным
афферентным путем она имеет собственное сенсорное звено.
4. Она не находится в антагонистических
отношениях с другими частями нервной системы.
5. Представляя истинно базовую
иннервацию, она обладает гораздо большей, чем симпатическая и парасимпатическая
нервная система, независимостью от ЦНС. —
6. Органы с разрушенными или с
выключенными с помощью ганглиоблокаторов метасимпатическими путями утрачивают
присущую им способность к координированной ритмической моторной и другим
функциям.
7. Метасимпатическая нервная
система имеет собственное медиаторное звено.
5.1.3.
Элементы эволюции автономной нервной системы
У беспозвоночных (аннелид и др.)
от соматической нервной системы отделяются нервные элементы, связанные с
кишечной трубкой. Они образуются из клеток глоточных узлов и формируют
самостоятельные ганглии. У членистоногих существует уже достаточно четко
дифференцированная автономная нервная система с узлами и нервными стволами,
идущими к мышцам желудка. У стрекозы, например, выделяют краниальный,
туловищный и каудальный отделы. Туловищный соответствует симпатической,
краниальный и каудальный — парасимпатической нервной системе позвоночных. У
низших черепных животных сформированных автономных ганглиев не обнаруживается,
хотя и имеются висцеральные ветви спинномозговых нервов.
У низкоорганизованных позвоночных,
таких как миноги, акулы, скаты, по ходу симпатических сплетений, снабжающих
пищеварительный канал и начинающихся от висцеральных ветвей спинномозговых
нервов, возникают ганглиозные скопления клеток. Они располагаются дорсальное
аорты соответственно каждой паре спинномозговых нервов. От ганглиев отходят
ветви к внутренним органам и сердцу. В толще органов эти ветви формируют
сплетения с заложенными в них ганглиозными клетками.
Таким образом, на этом этапе
эволюции возникает третья часть автономной нервной системы — метасимпатическая.
В ряду костистых рыб образуется
парный пограничный симпатический ствол со связями, характерными для высших
позвоночных. Краниально он заходит в голову, каудально достигает хвоста. В
дальнейшем в ходе эволюции строение автономной нервной системы усложняется,
отличаясь разнообразием у разных групп.
Симпатический ствол рептилий,
например, включает до 27 пар ганглиев с многочисленными связями, интрамуральные
сплетения внутренних органов хорошо дифференцированы, хотя число клеток в узлах
еще незначительно. У птиц преганглионарные волокна покидают спинной мозг в
составе вентральных корешков.
По мере филогенетического развития усложняется и клеточный
состав ганглиев. Если у низкоорганизованных позвоночных животных еще нет четких
специфических различий между нервными клетками, то у более высокоорганизованных
представителей класса рыб и особенно у млекопитающих имеется уже три их типа:
рецепторные, эффекторные и ассоциативные.
5.1.4. Различия в конструкции
автономной и соматической нервной системы
Первое различие — основное — заключается в расположении эффекторного
(двигательного) нейрона по отношению к ЦНС. В автономной нервной системе
эффекторная клетка находится за пределами спинного или головного мозга. В
соматической нервной системе нервные клетки, принимающие участие в
осуществлении рефлекса — вставочная и двигательная, — находятся внутри серого
вещества спинного мозга (см. рис. 5.1). Благодаря этому структурному отличию
возникают физиологические особенности, которые обусловливают несколько иное
протекание процессов в автономной нервной системе.
Второе различие относится к иннервации органов, снабжаемых волокнами
автономной и соматической нервной системы. Перерезка вентральных корешков
спинного мозга сопровождается полным перерождением всех эфферентных
соматических волокон и не затрагивает автономных в силу того, что ее
эффекторный нейрон вынесен в один из периферических ганглиев. Исполнительные
органы, в сущности, управляются импульсами только этого нейрона. Именно данное
обстоятельство и подчеркивает относительную автономию этого отдела нервной
системы.
Третье различие касается выхода нервных волокон. Соматические нервные
волокна покидают ствол мозга и спинной мозг сегментарно и перекрывают
иннервируемые области по меньшей мере трех смежных сегментов. Волокна
автономной нервной системы выходят из отстоящих друг от друга ограниченных
участков мозга: черепного, грудо—поясничного и крестцового. Волокна, выходящие
из грудо—поясничного отдела спинного мозга, и связанные с ними ганглии
называются симпатическими, а выходящие из черепного и крестцового — парасимпатическими.
Часть автономных рефлекторных дуг, локализованных в ганглиях стенок внутренних
органов (метасимпатическая нервная система), вообще не имеет прямых
выходов из ЦНС.
Четвертое различие относится к распределению нервных волокон на периферии.
Соматические волокна распределены строго сегментарно, волокна автономной
нервной системы этому правилу не подчиняются. Они иннервируют все органы без
исключения, а часть из них имеет двойную или даже тройную иннервацию —
симпатическую, парасимпатическую, метасимпатическую.
Помимо указанных имеются еще и сугубо морфологические
отличия. Волокна автономной нервной системы в большинстве своем лишены
миелиновой оболочки, тонкие, диаметр их не превышает 7 мкм, в то время как
соматические эфферентные волокна толстые, миелинизированные, достигающие в
диаметре 12—14 мкм. Отличия в строении связаны и с функциональными
особенностями — скоростью проведения, хронаксией. По тонким автономным
волокнам возбуждение распространяется значительно медленнее, чем по толстым
соматическим, и составляет соответственно 1—3 и 70—120 м/с. Для вызова ответной
реакции автономной нервной системы необходимо применять значительно большую
силу раздражения, так как ее волокна характеризуются большим рефракторным
периодом и большей хронаксией.
5.1.5.
Чувствительное звено дуги автономного рефлекса
Рецепторы висцеральных органов (интероцепторы)
по строению делятся на свободные и несвободные. Свободные окончания не
окружены вспомогательными образованиями или специальными клетками. К их числу
относят наиболее просто устроенные кустиковидные рецепторы, концевые структуры
которых имеют вид петелек, колечек, пластинок, включающих скопление митохондрий
и специфических пузырьков. Несвободные окончания имеют форму клубочков,
покрытых снаружи капсулой, пластинчатых телец, у которых рецепторное окончание
заключено в капсулу из нескольких слоев клеток и межклеточного вещества.
В процессе эволюционного развития
происходит увеличение числа интероцепторов, постепенно усложняется их форма и,
вероятно, также более узкой становится специализация. Сейчас у высших
позвоночных обнаружены интероцепторы, способные воспринимать механические и
химические стимулы.
Одиночные механорецепторы.
разделяют на два типа: медленно и быстро адаптирующиеся. На
стационарное непрерывное воздействие типа растяжения и сдавливания и те, и
другие отвечают залпом разрядов, частота которых находится в прямой зависимости
от скорости нарастания стимула и его амплитуды. Позже у медленно адаптирующихся
интероцепторов частота следования импульсов снижается до некоторого постоянного
уровня и, флуктуируя около среднего значения, может сохраняться часами, пока
действует раздражитель. У быстро адаптирующихся интероцепторов возросшая в
ответ на применение раздражителя активность вскоре затухает, а затем вовсе
исчезает. Эти различия в поведении интероцепторов связаны с особенностями
строения их структур.
Механорецепторы часто называют барорецепторами,
хотя, строго говоря, это типичные механорецепторы, так как их возбуждение
связано не с давлением, а с растяжением сосудистой или кишечной стенки, которое
в конечном счете приводит к увеличению поверхности воспринимающей части
мембраны.
Многочисленные попытки обнаружения
субстрата химической чувствительности также завершились определением
рецепторных структур, реагирующих на изменение химической среды.
В зависимости от типа реакции хеморецепторы
разделяют на кислото— и щелочечувствительные. И те, и другие
реагируют преимущественно на высокие концентрации веществ. Основным фактором,
способным возбудить деятельность рецепторных структур, является исключительно
рН растворов кислот и щелочей.
Интероцепторы располагаются в
стенках практически всех висцеральных органов, таких как сосуды, сердце,
легкие, почки, мочевой пузырь, печень и др.
Большинство артериальных
рецепторов является медленно адаптирующимися. Их активность находится в
прямой зависимости от уровня постоянного давления. Учащение импульсации в них
происходит линейно, пока давление не достигает величины 140—180 мм рт. ст.
(18,8—24,0 кПа). Наиболее отчетливо деятельность рецепторных единиц проявляется
при нарастании давления в связи с сердечным циклом: в ответ на каждую систолу
возникают пачки разрядов, исчезающих в период диастолы. У аортальных
рецепторов систолическая вспышка отстает от второго зубца ЭКГ на 70—80 мс.
При различных функциональных состояниях, болезнях сердца и сосудов работа
рецепторов перестраивается. Сосудистые интероцепторы стимулируются и угнетаются
определенными химическими веществами.
Среди рецепторов предсердий
установлено наличие двух видов, отличающихся друг от друга характером генерации
разрядов и ответами на изменение гемодинамики. В стенках желудочков также
существует по меньшей мере два типа рецепторов: вентрикулярные и эпикардиальные.
Особенностью первых является наличие раннего систолического залпа импульсов
перед открытием аортальных клапанов.
Подъем давления в желудочке путем
пережатия аорты или легочной артерии ведет к значительному увеличению разряда.
В отличие от вентрикулярных рецепторы эпикарда не реагируют на подобные
манипуляции, не действует на них и гипоксия. Их активность состоит из
нерегулярных разрядов с частотой 1—5 имп/с без проявления сердечного ритма.
Отдельные поля этих рецепторов составляют около 1 см2.
Рецепторы легких по
локализации разделяются на рецепторы растяжения, связанные с гладкомышечными
элементами стенок дыхательных путей, рецепторы слизистой оболочки и бронхов и
рецепторы, расположенные вблизи капилляров альвеол (юкстакапиллярные).
Первые наделены, как правило,
сугубо механорецепторными свойствами. Они реагируют только на растяжение
легочной ткани и в соответствии с этим активируются лишь при вдохе. Рецепторы,
лежащие более поверхностно, наряду с этим возбуждаются и химическими стимулами.
Это так называемые ирритантные рецепторы. Их активность изменяется в
связи с объемом легких реагируя на вдох и выдох, а также асфиксию, действие CO2 и
раздражающих паров. Юкстакапиллярные рецепторы являются быстро адаптирующимися
и мало способными к реактивации. Первоначально их обозначали как специфические
рецепторы спадения.
Рецепторы почки и мочевого
пузыря также подразделяются на быстро и медленно адаптирующиеся. Первые
воспринимают колебания давления в мочевом пузыре. На наполнение органа и его
опорожнение реагируют медленно адаптирующиеся структуры, что, вероятно, имеет
значение для рефлекторной регуляции деятельности органа (см. разд. 13.14).
Особый тип рецепторов описан в мочеиспускательном канале (уретре). Они способны
возбуждаться исключительно током жидкости. Частота их разрядов находится в
зависимости от скорости протекания мочи. Эти рецепторы локализуются в основном
вблизи сфинктера мочеиспускательного канала.
В печени имеются различные
популяции специфических рецепторов: волюмо—, осмо—, натрий— и калийчувствительные.
Они имеют различное функциональное значение: участвуют в системной регуляции
объема крови через орган и служат информационным звеном волюморегулирующего
рефлекса.
Применяя дозированное воздействие
на рецепторы, а также оценивая характер активности у голодных и сытых животных,
удалось определить наличие и локализацию рецепторов в пищеводе, желудке,
кишке, преджелудках жвачных. Большинство чувствительных окончаний пищевода относится
к быстро адаптирующимся. Они активируются в момент перистальтического
сокращения после глотания. В желудке этот вид рецепторов располагается в
кардиальной части, на дне и в пилорической части. В тонкой кишке также имеется
большое число участков, в которых удается регистрировать активность таких
рецепторов. Их высокая плотность обнаружена в преджелудках жвачных животных.
Рецепторные поля одиночных волокон большие, площадью до 4—5 см2. В
них расположены высокочувствительные зоны, находящиеся преимущественно в центре
поля. Их механическое раздражение сопровождается ответами с максимальной
частотой разрядов.
Рецепторы слизистой оболочки реагируют на легкое статическое
касание или длительное давление, на растяжение стенок желудка или кишки. Помимо
этих в кишке обнаружена еще одна разновидность механорецепторов. Они
возбуждаются только током жидкости в просвете кишки.
Последовательное удаление отдельных слоев желудочной стенки
позволило установить, что спонтанно разряжающиеся рецепторы располагаются преимущественно
в мышечном слое. В зависимости от пространственной ориентации — лежат ли они
параллельно или поперек хода мышечных волокон — рецепторы могут работать либо
как детекторы длины, либо как детекторы напряжения. Под серозной оболочкой
тонкой кишки и в мышцах слизистой оболочки располагаются крайне быстро
адаптирующиеся механорецепторы типа телец Фатера—Пачини. Величина рецептивного
поля каждой такой единицы достигает 1 мм2.
В наиболее крупных симпатических ганглиях также находятся
рецепторные окончания, способные возбуждаться в результате непосредственного
действия раздражителя и косвенно — путем изменения уровня метаболизма.
Реакция рецептора на стимул составляет основу для всего
последующего анализа сенсорных влияний, который выполняет нервная система.
Именно от рецепторов зависят пределы чувствительности и диапазон воспринимаемых
воздействий.
Чувствительные окончания по своим анатомическим и
функциональным особенностям устроены так, что избирательно реагируют лишь на
один определенный вид энергии. С другой стороны, стимул, каким бы он ни был по
своей природе, — механическая деформация, химическое изменение, температурный
сдвиг — всегда вызывает один и тот же электрический сигнал, который играет роль
символа. Информация об интенсивности стимула во всех рецепторах передается частотой
импульсов. Соотношение между интенсивностью стимула и частотой импульсации
устанавливается в результате взаимодействия между стационарным генераторным
током в дендритах чувствительной клетки и сдвигами проводимости во время потенциала
действия. Немаловажно и то, что рецепторы не только воспринимают, но и
усиливают воспринимаемый сигнал.
Нервные волокна, по которым висцеральная сенсорная
информация передается в центры интеграции, подразделяются на группы А,
Б, С. В отличие от метамерности, с которой в спинной мозг вступают соматические
афференты, проводники висцеральной чувствительности имеют более сложное
полисегментарное распределение, что создает условия для мультипликации их
действия.
При вступлении в спинной мозг часть афферентных волокон
первично связывается с сегментарными интернейронами, которые, в свою очередь,
синаптически контактируют с промежуточными или эфферентными клетками того же
или близлежащих сегментов. Так образуется дуга автономного рефлекса
спинального уровня. Вторая часть афферентных волокон распределяется
следующим образом: толстые миелинизированные волокна занимают более медиальное
положение, немиелинизированные — латеральное. Восходящие ветви частично
заканчиваются в сером веществе вышележащих сегментов, частично достигают
дорсальной части продолговатого мозга, где и заканчиваются в ядрах дорсальных
столбов. Здесь расположены вторые нейроны афферентного пути, аксоны которых
формируют бульботаламический путь. Нисходящие ветви спускаются на 6—7 см и
образуют синаптические контакты с ассоциативными клетками нижележащих
сегментов.
5.1.6.
Ассоциативное (вставочное) звено
В дуге автономного рефлекса
переключение афферентных сигналов на ассоциативные и далее на эфферентные клетки
может происходить в периферических образованиях — в превертебральных и в
интрамуральных ганглиях, так называемых низших центрах, а также на спинальном и
бульбарном уровнях.
Независимо от источника и природы
(висцеральной и соматической) афферентные сигналы иногда активируют одни и те
же нейронные механизмы, не имеющие существенных отличий в разных отделах ЦНС,
начиная от более простого сегментарного уровня, куда стекается первичная
информация, и кончая более сложным — надсегментарным. Они вовлекаются в разной
степени в зависимости от интенсивности воздействия и включаются в разных
соотношениях. Основой для запуска различных нейронных систем является мощность
поступающего афферентного потока.
Спинальные механизмы. В спинном
мозгу существует специальный интернейронный аппарат, осуществляющий первичную
обработку афферентных сигналов. Этот аппарат соединен с афферентными клетками
боковых рогов спинного мозга. Такая организация может, вероятно, согласовывать
афферентные сигналы, поступающие одновременно от различных рецептивных зон.
Интеграция сигналов не ограничивается сегментарным спинномозговым уровнем и в
определенных условиях может осуществляться внутрицентральными системами и
уровнями более высокого порядка.
Основные данные о сегментарных
вставочных нейронах получены на грудных и поясничных сегментах. Они
свидетельствуют об их четкой структурной дифференциации. Вставочные нейроны с
синаптическим входом от афферентных путей внутренних органов локализуются
главным образом в глубоких участках серого вещества (V пластина, по Рекседу).
Сегментарный механизм
взаимодействия чувствительных клеток автономной нервной системы и
преганглионарных симпатических нейронов не может быть целиком сведен к функции
универсального «интегрирующего» вставочного нейрона. Такую передачу может
осуществлять только соответственно организованный ансамбль клеток. Среди них
находятся, например, вставочные нейроны, которые способны к суммации
висцеральных и соматических влияний, что приводит в конечном счете к взаимному
облегчению прохождения каждого из них. Помимо того, имеются вставочные клетки,
в которых афферентные автономные и соматические влияния вызывают ответы
смешанного типа (ВПСП—ТПСП), т. е. помимо деполяризационных возможно развитие и
гиперполяризационных изменений. Это означает, что при определенных условиях
один из ответов может подавляться, в связи с чем пресинаптической
гиперполяризации отводят роль облегчающего фактора в прохождении, например,
болевых сигналов, хотя такая точка зрения не является общепринятой.
Простейшие дуги автономных
рефлексов спинального уровня не имеют прямых переключении с
афферентных волокон на преганглионарные нейроны. Как установлено, самый
короткий путь между ними включает два синапса. Для сравнения — в соматической
дуге моносинаптического рефлекса растяжения существует всего одно переключение
(см. разд. 1.1.9).
Следовательно, в дуге автономного
рефлекса между чувствительным и постганглионарным нейронами расположено не
менее трех синапсов, два из которых находятся в сером веществе спинного мозга,
а один — в периферическом автономном ганглии.
|
|
|
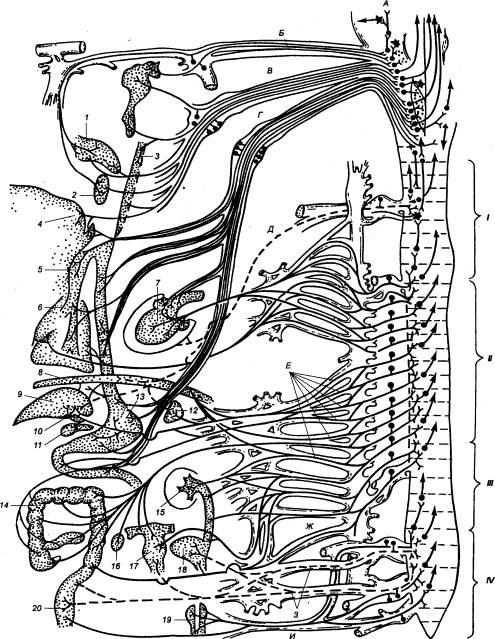
Рис. 5.7 Распределение
висцеральных чувствительных волокон в сегментах спинного мозга (по
Черниговскому) Слева
изображены волокна, отходящие от внутренних органов, справа — сегменты спинного
мозга и входящие в них волокна. I — шейный, II — грудной, III —
поясничный, IV — крестцовый отделы спинного мозга: А — медиальная петля, В
— лицевой нерв, В — языкоглоточный нерв, Г — блуждающий нерв, Д — диафрагмальный нерв, Е — внутренностные нервы, Ж —
подчревный нерв, 3 — тазовые
внутренностные нервы, И —
половой нерв; 1 — язычок, 2 — нёбо, 3 — глотка, 4 — язык, 5 — трахея, 6 — легкие, 7 — сердце, 8 — диафрагма, 9 — печень, 10 — жёлчный пузырь, 11
— желудок, 12 — селезенка,
13 — поджелудочная железа, 14 — тонкая кишка, 15 — почка, 16 — яичник, 17 —
матка, 18 — мочевой пузырь, 19 — мочеиспускательный канал, 20 — толстая кишка. |
Одним из возможных медиаторов пресинаптического
взаимодействия в спинном мозгу считают ГАМК, в то же время не подлежит
сомнению и участие норадреналина (см. разд. 5.2.2) — одного из посредников
нисходящих супраспинальных влияний на интернейронный аппарат спинного мозга.
Вклад в осуществление висцеро—висцерального взаимодействия вносит и механизм
возвратного торможения, что особенно отчетливо выражено у крестцовых
парасимпатических нейронов. Вероятно, аналогичный принцип постсинаптического
регулирования выражен и у преганглионарных симпатических клеток.
Результаты прямого электрофизиологического изучения ассоциативного
звена спинального уровня (преганглионарных нейронов) указывают на
исключительную сложность его организации и функциональной ориентации.
Существование различных собственных выходных элементов, возможное присутствие в
связи с этим специализированных возбуждающих интернейронов, тормозных
механизмов (например, возвратного торможения) и нисходящих симпатоактивирующих
путей делают вполне оправданным введение В. П. Лебедевым в обозначения этой
системы специального понятия симпатический механизм спинного мозга.
Супраспинальные механизмы. Дальнейшая обработка афферентных сигналов
висцеральной чувствительности, поступающих по проводящим путям спинного мозга,
происходит в ретикулярных образованиях заднего мозга, коре мозжечка,
вестибулярных ядрах и других структурах. Помимо того, здесь взаимодействуют
сигналы, приходящие и по афферентным волокнам черепных нервов (рис. 5.7).
Взаимодействия афферентных висцеральных и соматических
сигналов активируют ретикулярные нейроны, которые посылают нисходящие сигналы к
спинальным соматическим и преганглионарным нейронам дуги автономного рефлекса.
Эти нисходящие влияния могут носить тормозный или возбуждающий характер. У
части ретикулярных нейронов висцеральная афферентная волна приводит к
генерализации возбуждающих реакций. Все это указывает на существование здесь
сложных процессов переработки информации, что может проявляться, например, в
виде длительной неспособности к реактивации при повторных раздражениях (П. Г.
Костюк).
По внутрицентральным восходящим путям висцеральные потоки
достигают среднего мозга, ретикулярная формация которого является существенной
инстанцией на пути к ассоциативной области коры больших полушарий (рис. 5.8.).
|
|
|
Рис. 5.8 Схема проведения висцеральных сигналов по лемнисковым
и экстралемнисковы/м системам (по Черниговскому) |
Проекции афферентных путей висцеральных нервов диффузно
распределяются по всей ретикулярной формации за исключением рострального отдела
(В. Н. Черниговский). Особенностью представительства является то, что в одной и
той же точке регистрируются ответы не только от висцеральных, но и соматических
чувствительных окончаний. Полная утеря модальной принадлежности происходит еще
на подступах к ретикулярной формации, т. е. в формирующих восходящие пути
системах промежуточных нейронов.
Для каждой из афферентных систем, за исключением
обонятельной, в промежуточном мозгу имеется специальное переключающее первичное
«реле» с высокой степенью дифференциации и плотностью представительства. На
этом уровне могут осуществляться не только различные переключения на
висцеральные рефлекторные системы, но и взаимодействия между различными
рефлекторными сигналами. К структурам промежуточного мозга, осуществляющим эту
функцию, относятся специфические и неспецифические ядра таламуса, а также
гипоталамические структуры (см. разд. 3.7.3).
Висцеральные афферентные сигналы, поступающие в спинной
мозг, могут достигать таламических структур по двум системам: задние столбы—
медиальная петля, передний и боковой спинно—таламические пути. Согласно
классическим представлениям, специфические проекционные пути включают три
нейрона. Аксон первого, располагающегося в спинальном или одном из автономных
ганглиев, достигает тонкого и клиновидного ядер продолговатого мозга, где
переключается на второй нейрон. Аксон второго нейрона всех специфических
проводящих путей оканчивается на клетках противоположной стороны вентрального
заднелатерального ядра таламуса. Здесь располагается третий нейрон. Его аксон
проецируется в соматосенсорные области коры головного мозга.
Зона представительства основного коллектора чувствительных
путей брюшной полости — чревного нерва — располагается в латеральной части зад
невентрального ядра таламуса. Несколько иначе представлен блуждающий нерв. Его
ответы регистрируются в медиальной зоне того же ядра. Характерно, что в зоне
представительства блуждающего нерва не удается зарегистрировать ответов
чревного нерва. Вместе с тем области представительства каждого из этих нервов
перекрываются проекциями соматических афферентов: внутренностного — от
туловища, блуждающего — от плечевого пояса, тазовых — от задних конечностей.
В этом отношении они близки к проекциям тех же нервов в
соматосенсорных областях коры головного мозга. Это означает, что в обоих
проекционных ядрах таламуса происходит взаимодействие информации, поступающей
по висцеральным и соматическим афферентным путям. В результате происходит
утрата специфичности информации, и к корковым клеткам направляются уже
интегрированные сигналы. Объединение сигналов висцеральной и соматической
природы, помимо того, происходит и в самой коре мозга. Какого—либо специального
места для переключения сугубо висцеральных сигналов в коре головного мозга не
существует.
При взаимодействии конвергирующих окончаний висцеральных и соматических
афферентов на одних и тех же нейронах сигналы, идущие по соматическим путям,
блокируют доступ висцеральной информации к конвергирующей клетке. Такое
положение послужило основанием считать (Р. А. Дуринян), что в релейных ядрах
таламуса ограничивается поступление висцеральной информации в корковые
структуры и их нейронные системы обслуживают процессы, требующие лишь
непременного коркового контроля. Эти соотношения могут меняться в зависимости
от ситуации. Иногда при значительном усилении висцеральной афферентации зона
представительства висцерального нерва расширяется, занимаются поля перекрытия,
в результате чего происходит ограничение передачи соматических импульсов.
Переработка афферентной сигнализации может происходить не только в таламических
ядрах, но и в заднелатеральных участках гипоталамуса (О. Г. Баклаваджян).
Кортикальное представительство. В организации проекций афферентных
систем в коре больших полушарий выделяют два типа представительства:
первичное и вторичное. Первичное связано с локальным
распределением в одной области коры преимущественно одного типа чувствительных
связей. При этом ответы возникают при раздражении любого звена афферентной
системы. Особенностью вторичного представительства является диффузное
распределение проекций афферентных систем разной чувствительности. Они
регистрируются за пределами первичных проекционных областей (В. Н.
Черниговский).
Чувствительные импульсы, идущие от органов брюшной полости
по внутренностным нервам, проецируются в определенные области коры, проявляясь
контра— и ипсилатерально в I и II соматосенсорных областях (рис. 5.9). Здесь
также представлена и соматическая чувствительность. На этом основании считают,
что объединение сигналов висцеральной и соматической природы происходит не
только в таламических ядрах, но и в коре мозга. В пользу такого представления
свидетельствуют и наблюдения о взаимной окклюзии ответов при раздражении
соматических и висцеральных афферентов, зарегистрированные на отдельных
нейронах.
Импульсы, идущие от шейной, грудной и поддиафрагмальной
частей блуждающего нерва, регистрируются в глазничной извилине, между концом
коронарной и ринальной извилин, между глазничной и носовой бороздами, а также в
поясной извилине медиальной поверхности мозга. В орбитальной области коры имеется
перекрытие зон блуждающих и внутренностных нервов, однако фокусы их
максимальной активности в значительной мере разобщены.
Рассмотрение коркового представительства тазового нерва
указывает на то, что он обнаруживает изолированные проекции в двух областях.
Одна проекция располагается в средней части крестовидной борозды и частично
заходит на медиальную поверхность полушария, другая — находится во второй
соматической зоне.
Представительства функционально близких висцеральных систем
находятся и в близко расположенных областях коры. Так, зоны брыжеечных,
селезеночных и чревных нервов перекрываются зонами представительства
блуждающего нерва, что является основой для тонкой и надежной координации
процессов,
|
|
|
Рис. 5.9 Карта представительства
афферентных систем внутренних органов в коре больших полушарий 1—6,
13—16, 19 — блуждающий нерв (1—4,
19 — шейный отдел нерва, 5—6
— брюшной отдел нерва), 7,8 —
тазовый нерв, 9—10 —
внутренностный нерв, 11—12 — рецепторы
сердца и венечных (коронарных сосудов), 17, 18 — гортанный и языкоглоточный нервы. |
осуществляемых корой. Последнее свойство подтверждается
существованием множественного представительства висцеральных систем в коре
мозга. И в самом деле, сердце, пищеварительный тракт — непарные органы; в
случае нарушения их работа не может компенсироваться другим органом; это
обязывает иметь двойное представительство в коре.
Локализация в коре проекций висцеральных органов и систем в
ближайшем соседстве с соматическими и ассоциативными обеспечивает, видимо,
появление необходимых висцеральных рефлекторных реакций. Сигнализация,
следующая по висцеральным афферентным путям и достигающая первичных и вторичных
зон представительства, вместе с соматической чувствительностью участвует в
формирований механизмов целенаправленного поведения.
Несмотря на сложность многоступенчатой организации, основа
механизма межнейронного взаимодействия на всех уровнях одна и та же —
синаптическая конвергенция на центральных клетках сигналов различной природы
(висцеральной и соматической), их оценка аппаратом суммации и на основании
результатов оценки — генерация нового сигнала.
Как установлено в последние годы, различия между
механизмами, включающимися на различных уровнях — спинальном, супраспинальном,
корковом, — не принципиальны и заключаются в основном в количественных
особенностях конвергенции.
Преганглионарные симпатические волокна, выходящие из боковых
рогов серого вещества спинного мозга в составе белых соединительных ветвей (см.
рис. 5.1), достигают паравертебральных симпатических ганглиев, где часть их
синаптически контактирует с эфферентными нейронами. Другие волокна без
синаптического переключения в этих узлах следуют далее и вступают в
синаптический контакт уже в превертебральных ганглиях, таких как комплекс узлов
чревного сплетения. Исключение из правил составляет часть волокон, следующих в
составе внутренностного нерва к надпочечнику. Эти преганглионарные волокна
проникают непосредственно в мозговой слой железы, выполняющий в данном случае
роль своеобразного постганглионарного звена.
Преганглионарные нейроны получают входы от периферических
висцеральных и соматических чувствительных клеток, а также по нисходящим путям
от высших центров.
Преганглионарные волокна различаются по возбудимости,
скорости проведения возбуждения и другим функциональным свойствам. Наиболее
тонкие и низковозбудимые, как, например, симпатические дилататоры (расширители)
направляются к сосудам скелетных мышц. Самый низкий порог возбуждения у
волокон, идущих к мигательной перепонке и мышце зрачка.
Подходя к эффекторным клеткам, преганглионарное волокно
теряет миелин и, ветвясь, образует на теле и отростках постганглионарных клеток
синаптические контакты. Преганглионарных волокон значительно меньше, чем
постганглионарных нейронов. Так, к верхнему шейному симпатическому ганглию у
человека подходит примерно 10 тыс. Преганглионарных волокон, которые
оканчиваются на миллионе постганглионарных клеток узла.
Таким образом, одно преганглионарное волокно контактирует не
менее чем с сотней постганглионарных нейронов. У кошки это соотношение
составляет 1 : 32. На одном и том же постганглионарном нейроне может
конвергировать несколько Преганглионарных волокон. Такие широкие дивергенция и
конвергенция обеспечивают высокую надежность проведения возбуждения в ганглии.
В интеграции сигналов в этих низших рефлекторных центрах определенную роль
играет пространственная и временная суммация постсинаптических потенциалов,
поскольку одиночные импульсы, обычно поступающие по преганглионарным аксонам,
не способны вызвать в постганглионарных нейронах сверхпороговые
постсинаптические потенциалы.
В парасимпатических узлах такая конвергенция практически
отсутствует. Каждое преганглионарное волокно контактирует только с одной—двумя
постганглионарными клетками. Этим объясняется то, что парасимпатическая
стимуляция завершается, как правило, менее распространенной, чем симпатическая,
ответной реакцией.
Преганглионарные волокна белых ветвей большей частью
спонтанно не активны. Их стимуляция сопровождается появлением характерных
сдвигов мембранных потенциалов и возникновением ПД. Вместе с тем
Преганглионарные нейроны, принимающие участие в вазомоторной регуляции и
возбуждающиеся сигналами нисходящих симпатоактивирующих путей, являются
спонтанно активными. Сумма разрядов нескольких волокон выявляет наличие ритмов,
присущих целым нервным стволам бодрствующих животных — ритму пульсовых толчков
и дыхательных движений. Эта активность отражает тонические влияния, например,
вазомоторного центра. Тоническая и вызванная рефлекторная активность
опосредуется, по—видимому, различными клетками.
Взаимодействуя с метасимпатической нервной системой,
Преганглионарные парасимпатические и постганглионарные симпатические волокна
(см. рис. 5.1 и 5.4) модулируют активность исполнительного висцерального
органа. При этом допускается, что парасимпатические волокна способны возбуждать
не только эфферентные нейроны метасимпатической нервной системы, активирующие,
например, моторную деятельность кишки, но и тормозные клетки. Симпатические же
влияния наряду со слабо выраженным прямым действием на гладкие мышцы органа
вызывают возникновение тормозных процессов в холинергических нейронах местных
метасимпатических сетей, а также подавляют выход медиатора из преганглионарных
парасимпатических волокон. Следовательно, модулируя приток импульсов по
парасимпатическим путям, симпатическая нервная система тем самым оказывает
регулирующее влияние на моторную активность таких органов, как кишка и мочевой
пузырь.
Эфферентное звено дуги автономного рефлекса представляет
собой нервную клетку, мигрировавшую из спинного мозга в один из периферийных
ганглиев (рис. 5.10). По длине аксонов эти клетки разделяют на короткие
и длинные, по характеру ветвления терминалей — на густо и равномерно
сетевые. Постганглионарные волокна иннервируют большинство органов и
тканей. Преобладающей формой межнейронных связей, посредством которых
осуществляются контакты между преганглионарным волокном и эффекторным нейроном,
являются так называемые перицеллюлярные аппараты.
Потенциал покоя эффекторного нейрона дуги автономного
рефлекса незначительно отличается от потенциала покоя соматического
мотонейрона. Одиночная или кратковременная стимуляция преганглионарных волокон
изолированного ганглия сопровождается сложной последовательностью медленных
потенциалов: локальный отрицательный потенциал — 0—волна,
переходящий в положительную П—волну, которая сменяется поздней
отрицательной ПО—волной. О— и ПО—волны являются результатом возбуждения
преганглионарных волокон, контактирующих с ганглионарной клеткой, 0—волна
представляет ВПСП, возникающий в холинергических синапсах, ПО — поздний ВПСП.
|
|
|
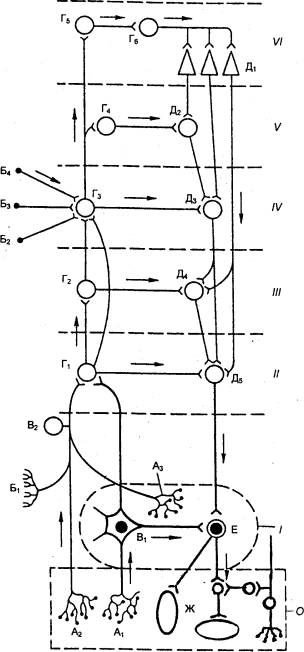
Рис. 5.10 Основные связи автономной нервной системы (по Семенову с
изменениями) О —
висцеральный орган; справа внизу изображен функциональный модуль
метасимпатической нервной системы, локализующийся в интрамуральных ганглиях
стенок полых висцеральных органов; I —
автономный (околопозвоночный или предпозвоночный) узел; II — сегмент спинного
мозга или ствола мозга; III — надсегментарные центры продолговатого мозга; IV
— промежуточный мозг; V — ядра основания мозга; VI — кора больших полушарий
головного мозга; A1 и A2 — рецепторы внутренних
органов; A3—
рецептор автономного нервного узла; Б1 — экстероцептор; Б2—Б4 — проводящие
пути сенсорных систем; B1 —
чувствительный нейрон автономного нервного узла; B2 — чувствительные нейроны спинномозговых и черепно—мозговых
узлов; Г1—Г4 — ассоциативные ядра разных отделов мозга; Г5 — клетки афферентных
зон коры головного мозга; Г6 — ассоциативные
клетки; Д1— пирамидные клетки коры головного мозга; Д2 — ядра основания. Д3 —
ядра гипоталамуса; Д4 — клетки ретикулярной формации продолговатого мозга; Д5
— ядра автономной нервной системы в стволе мозга и сегментах спинного мозга;
Е — эфферентный нейрон дуги автономного рефлекса; Ж — эфферентные клетки
стенки висцерального органа. Стрелками обозначено распространение
возбуждения. |
Появление П—волны связывают с возбуждением
преганглионарных волокон, оканчивающихся на хромаффинных клетках ганглия.
Функциональное значение длительных синаптических процессов в ганглии остается
неясным.
Возникновение медленных
деполяризующих и гиперполяризующих постсинаптических потенциалов зависит в
основном от мускариноподобного действия ацетилхолина. Однако
гиперполяризация опосредована вставочными адренергическими клетками
симпатических ганглиев, участвуя таким образом в регуляции возбудимости
постганглионарных клеток. Все это служит еще .одним из свидетельств в пользу
того, что ганглии автономной нервной системы являются низшими интегративными
центрами.
Эфферентный нейрон может иметь
несколько входов. Он обладает рядом морфологических и функциональных свойств,
которые позволяют интегрировать входные сигналы и образовывать модифицированный
выходной сигнал. Основной вход —возбуждающий, холинергический — представлен
преганглионарными волокнами. Следующий вход сугубо периферического
происхождения.
Клеточные тела этих чувствительных
нейронов могут располагаться либо в самом ганглии, в непосредственной близости
от эфферентного нейрона, либо в ганглиях стенок внутренних органов, т. е. имеют
метасимпатическую природу. Благодаря этим местным рефлекторным дугам в
эфферентном нейроне поддерживается необходимый уровень спонтанной активности и
сохранение рефлекторной деятельности при децентрализации узла. Величина
синаптической задержки — время между возбуждением пресинаптической терминали и
образованием ВПСП в одиночном нейроне — составляет 1,5—2 мс, длительность ВПСП
20—50 мс, пороговая амплитуда ВПСП для разряда 8—25 мВ, длительность разряда
1,5—3 мс.
Часть эфферентных нейронов
спонтанно активна. В некоторых из них ритм и паттерн разряда совпадают с
показателями активности преганглионарных
|
|
|
Рис.
5.11 Симпатическая нейрональная
активность в момент перистальтического сокращения (А. Б) и расслабления (В)
участка ободочной кишки на А, Б фазы: а —
расслабления, б — сокращения,
в — спада; на В фазы: а — спада, б — расслабления, в — сокращения; 1 — нейрограмма, 2 —
энтерограмма, 3 — пневмограмма, 4 —
ЭКГ. |
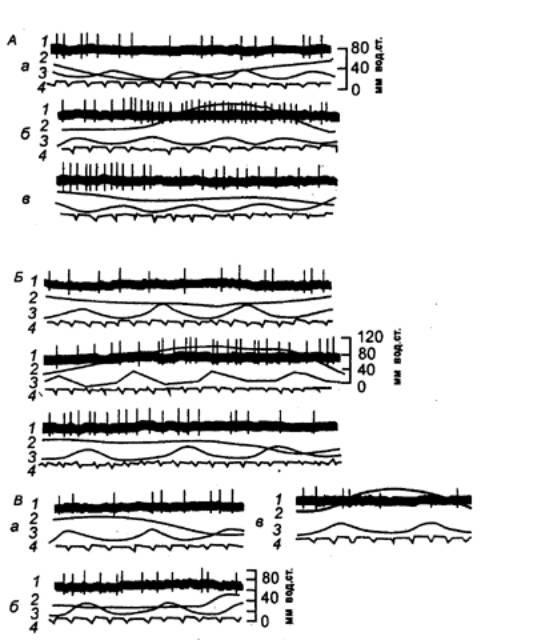
волокон
или отражают приток по висцеральным волокнам местных рефлекторных дуг. В большинстве
отмечается хорошая корреляция «входов» и «выходов». Фоновая активность
эфферентных нейронов низкочастотна, разряды клеток могут появляться в момент
сердечного толчка, фазы вдоха или (как, например, у брыжеечного ганглия) еще и
синхронно с перистальтическими движениями кишки. При этом выделяют две группы
клеток: одни разряжаются в момент сокращения, другие — в момент расслабления
кишечной петли (рис. 5.11).
В иннервируемую гладкую мышцу постганглионарные
адренергические волокна вступают в виде пучков, включающих до 100 аксонов.
Каждый аксон составляет около нескольких сотен микроэлементов и оканчивается
россыпью то утолщающихся, то утончающихся веточек, образуя варикозные
расширения.
В
автономной нервной системе существует три вида синоптической передачи:
электрическая, химическая и смешанная.
Электрические синапсы располагаются преимущественно там, где необходима быстрая
передача возбуждения (например, ресничный ганглий птиц). В них не бывает
синаптической задержки и электрический сигнал проходит большей частью в обоих
направлениях. От химического синапса электрический отличается
симметричностью и тесным контактом мембран. В местах контакта суженная
синаптическая щель перекрыта тонкими канальцами, которые создают возможность
быстрого перемещения ионов между клетками. В пре— и постсинаптических
образованиях часто встречаются синаптические пузырьки, которые могут находиться
одновременно в обеих структурах. Полагают, что они переносят помимо медиаторов,
и трофические вещества.
Существуют также смешанные синапсы (например,
чашечкообразные окончания в ресничном ганглии цыпленка), в которых
электрический контакт занимает только часть площади синапса, остальное же по
морфологическим показателям и функциональным свойствам представляет собой
типичный химический синапс.
Синапсы с химической передачей характеризуются рядом
морфологических особенностей. Эти особенности и механизмы химической передачи в
автономной нервной системе принципиально те же, что и в нервно—мышечной
концевой пластинке (см. разд. 1.1.7). Однако в автономной нервной системе пре—
и постсинаптические образования значительно разнообразнее. Это нейроны, гладкомышечные,
железистые и миокардиальные клетки. Широко варьирует также плотность автономной
иннервации разных гладкомышечных органов.
На органах с обильной иннервацией
находятся прямые нервно—мышечные соединения, в которых расстояние между
расширением аксона и мембраной гладкомышечной клетки составляет около 20 нм. К
числу таких органов относятся семявыносящий проток и ресничная мышца. Здесь
гладкомышечные клетки полностью подчиняются нервному контролю, и катехоламины
крови на них не оказывают действия. У большинства же кровеносных сосудов
адренергическая иннервация захватывает только адвентициальную оболочку и частично
прилежащие узкие участки средней оболочки.
Следовательно, расстояние между
расширением аксона и гладкомышечными волокнами составляет более 80 нм. В
результате такой организации на гладкую мышцу сосудов значительное влияние
оказывают катехоламины крови, легко диффундирующие к ней через интиму. При этом
следует отметить, что общая длина конечных ветвей адренергического нейрона
достигает 30 см, число расширений составляет 250—300 на 1 мм.
Закономерности функционирования
синоптических аппаратов с химической передачей возбуждения были сформулированы
относительно давно. Первый принцип заключался в том, что нейрон со всеми
отростками выделяет один и тот же медиатор (принцип Дейла). Согласно второму
принципу, действие каждого медиатора на нейрон или эффектор определяется
природой рецепторов постсинаптической мембраны.
Согласно принципу Дейла, каждый
нейрон для передачи своей специфической информации использует только один
медиатор. Однако, как оказалось позже, наряду с основным в этом нейроне могут
присутствовать также другие передатчики и вещества, участвующие в их синтезе
(см. разд. 1.1.7).
Сейчас в автономной нервной
системе насчитывается более десятка типов нервных клеток, которые продуцируют
разные медиаторы — ацетилхолин, норадреналин, серотонин и другие биогенные
амины, АТФ и аминокислоты. В связи с этим их называют холинергическими,
адренергическими, серотонинергическими, пуринергическими и т. п. нейронами.
Столько же типов существует гипоталамических нейронов, продуцирующих
нейрогормоны. Согласно Д. А. Сахарову, главной причиной разнообразия
медиаторных механизмов является множественность происхождения (полигенез)
самих нейронов.
Ацетилхолин — это вещество,
которое в полной мере отвечает требованиям, позволяющим считать его химическим посредником
передачи возбуждения:
локализация ацетилхолина и
участвующего в его синтезе фермента в пресинаптической терминали и
соответствующей нервной клетке, его высвобождение при возбуждении; выделение
при стимуляции нервных окончаний; наличие в постсинаптической мембране
высокочувствительных химических структур (рецепторов), а также ферментативных
систем для инактивации вещества; одинаковое со стимуляцией пресинаптических
структур действие некоторых фармакологических веществ, искусственной аппликации
или подведения медиатора к ганглионарной клетке посредством ионофореза.
Присутствие ацетилхолина как
медиатора было прежде всего установлено в сердце лягушки, затем в сердце
млекопитающих и позже во многих периферических и центральных структурах. Сейчас
считают, что ацетилхолин высвобождается в окончаниях всех преганглионарных (и
симпатических, и
|
|
|
Рис.
5.12 Основные трансмиттеры в
химической передаче возбуждения в периферической нервной системе
млекопитающих АХ — ацетилхолин,
АД — адреналин, НА —
норадреналин; AX1
блокируется ядом кураре, АХ2 блокируется ганглиоблокаторами (например,
гексонием), AX3
блокируется атропином. |
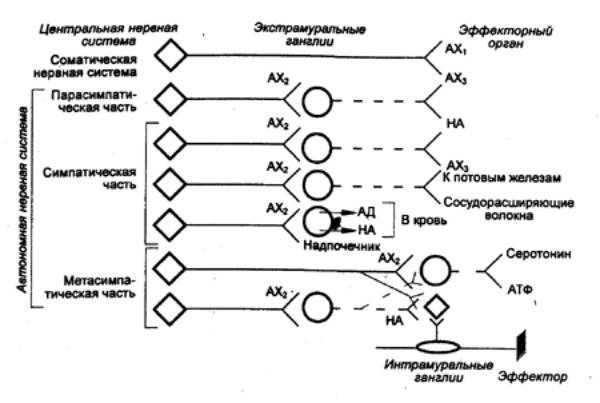
парасимпатических) нейронов и большинства постганглионарных
парасимпатических нейронов. Кроме того, часть постганглионарных симпатических
нейронов, идущих к потовым железам, и, по—видимому, симпатические нейроны,
вызывающие расширение сосудов скелетных мышц, также осуществляют передачу с
помощью ацетилхолина (рис. 5.12).
Само действие ацетилхолина может быть воспроизведено с
помощью фармакологических препаратов. Так, никотин вызывает подобный
ацетилхолину эффект при действии на постсинаптическую мембрану
постганглионарного нейрона, сложные эфиры холина, пилокарпин и токсин
мухомора мускарин — на мембрану эффекторного органа. Такое различие в
вызываемых реакциях послужило основанием к выделению двух типов
холинорецепторов: никотинового (Н—холинорецептор) и мускаринового (М—холинорецептор).
Фармакологические препараты, которые оказывают на
Эффекторный орган действие, аналогичное постганглионарному парасимпатическому
нейрону, получили название парасимпатомиметиков. Существуют также
вещества, избирательно блокирующие функцию каждого из холинорецепторов, названные
ганглиоблокаторами (ганглиолитиками).
Н—холинорецептор блокируется аммониевыми соединениями. М—холинорецептор
выключается атропином и скополамином. Ганглиоблокаторы,
выключающие или ослабляющие действие ацетилхолина на исполнительный орган,
называются парасимпатолитиками. Примером последних может служить атропин.
Помимо препаратов, действующих прямо или косвенно на
холинорецепторные структуры, большое распространение получили вещества,
оказывающие влияние на процессы синтеза и высвобождения ацетилхолина. Это
прежде всего гемихолиний и ботулиновый токсин. Первый нарушает
транспорт холина и тормозит синтез медиатора, второй — блокирует высвобождение
ацетилхолина.
Ацетилхолин кроме медиаторной роли обладает и общебиологическим
действием. Наиболее чувствительна к нему сердечно—сосудистая система. Он
также усиливает легочную вентиляцию, сокращает бронхиальную мускулатуру и
понижает бронхиальную секрецию, вызывает усиленную желудочно—кишечную
перистальтику, активируя одновременно секрецию желудочных, кишечных и
поджелудочных желез. Из—за кратковременного эффекта сам ацетилхолин применяют
редко, чаще используют метахолин и карбамилхолин. Их действие более
продолжительно и более сильно.
5.2.2. Норадреналин и
адреналин
После того как О. Лёви (1921)
доказал, что в результате раздражения симпатического нерва сердца в нем
высвобождается ускоряющее сердечную деятельность вещество, началось изучение
химической структуры и распределения этого вещества. Итогом поисков явилось
предположение о существовании двух типов симпатинов: возбуждающего и
тормозящего. Позже было установлено, что это норадреналин и адреналин.
Норадреналин
отличается от адреналина отсутствием метилового радикала в боковой цепи
(см. рис. 1.41). Отсюда возникло и его название — аббревиатура немецкого Nitrogen ohne Radical — NOR. Норадреналин является медиатором в
постганглионарных симпатических окончаниях сосудов сердца, печени, селезенки.
Его содержание достигает почти 10% от общего количества катехоламинов в областях
симпатической иннервации. Особенно богаты им органы, содержащие хромаффинную
ткань. В мозговом слое надпочечников, клетки которого гомологичны
постганглионарному симпатическому нейрону, содержание норадреналина составляет
около 20%, остальное приходится на долю адреналина. Наряду с тем и другим в
тканях содержится еще третий естественный катехоламин — дофамин.
Соотношения между адреналином, норадреналином и дофамином варьируют в
зависимости от вида и возраста животного.
Вещества, воспроизводящие эффекты
раздражения постганглионарных симпатических нейронов, получили название симпатомиметических
веществ; напротив, вещества, предотвращающие это действие, — симпатолитических.
Попадая на постсинаптическую
мембрану, норадреналин взаимодействует с ее специфическими химическими
компонентами — адренорецепторами. С адренорецептором может
взаимодействовать не только медиатор, но и аналогичные фармакологические
средства, называемые адреномиметическими, а также средства типа
дигидроэрготомина, дигидроэрготоксина, пропранолола, блокирующие
адренорецепторы.
В соответствии с чувствительностью
к различным катехоламинам (обычно к адреналину, норадреналину, изопротеренолу)
адренорецепторы делят на α—адренорецепторы и β—адренорецепторы.
Действие вещества считается α—адренергическим, если при его
воспроизведении эффективность убывает в последовательности норадреналин—адреналин—изопротеренол,
а также избирательно блокируется (β—адреноблокаторами. Если же
воспроизведение действия будет происходить в обратной последовательности:
изопротеренол—адреналин—норадреналин — и этот эффект предотвращается
специфическими блокаторами, то в этом случае имеет место β—адренергический
эффект.
В большинстве висцеральных
органов, реагирующих на катехоламины, находятся оба вида адренорецепторов и
результаты их возбуждения бывают, как правило, противоположными. Например, в
гладкой мышце артерий скелетных мышц содержатся α— и (β—адренорецепторы.
Возбуждение α—адренорецепторов приводит к сужению артериол, а возбуждение
β—адренорецепторов — к их расширению. В кишке также имеются оба вида
адренорецепторов (см. разд. 11.1.3), однако воздействие на те и другие вызывает
торможение активности гладкой мышцы. В обычных жизненных условиях реакция
органа на адреналин и норадреналин, выделяющиеся нервными окончаниями или
приносимые кровью, зависит от преобладания α— или (β—адренергического
действия (табл. 5.1).
В других висцеральных органах
находится только один вид адренорецепторов. В сердце и бронхах нет α—адренорецепторов,
и здесь норадреналин
Та
б л и ц а 5.1
Показатели
изменения функций различных органов при стимуляции симпатических и
парасимпатических нервов
|
Орган или система |
Симпатические нервы и
адренорецепторы |
Парасимпатические нервы |
|
|
Пищеварительный тракт: |
|||
|
продольные и циркулярные мышцы |
ослабление моторики |
α,β |
усиление моторики |
|
сфинктеры |
сокращение |
α |
расслабление |
|
Мочевой пузырь: |
|||
|
мышца, выталкивающая мочу |
расслабление |
β |
сокращение |
|
внутренний сфинктер |
сокращение |
α |
— |
|
Бронхиальные мышцы |
расслабление |
β |
сокращение |
|
Внутриглазные гладкие мышцы: |
|
||
|
дилататор зрачка |
сокращение |
α |
— |
|
сфинктер зрачка |
— |
— |
сокращение |
|
ресничная мышца |
расслабление |
β |
—«— |
|
Пиломоторные мышцы. |
сокращение |
α |
— |
|
Половые органы: |
|||
|
семенные пузырьки |
сокращение |
α |
— |
|
семявыносящий проток |
—«— |
—«— |
— |
|
матка в зависимости от вида |
—«— |
—«— |
|
|
матка в зависимости от
гормонального фона |
расслабление |
β |
— |
|
Сердце: |
|||
|
ритм |
ускорение |
β |
замедление |
|
сила сокращения |
увеличение |
—«— |
ослабление |
|
Кровеносные сосуды: |
|||
|
артерии кожи |
сужение |
— |
— |
|
артерии брюшной полости |
—«— |
— |
— |
|
артерии скелетных мышц |
—«— |
— |
— |
|
артерии венечные (коронарные) |
сужение, расширение |
α |
— |
|
сосуды мозга |
сужение |
—«— |
расширение (?) |
|
артерии половых органов |
——«— |
—«— |
расширение |
|
вены |
—«— |
—«— |
— |
|
Экзокринные железы: |
|||
|
слюнные |
секреция |
α |
секреция |
|
слёзные |
— |
|
—«— |
|
пищеварительные |
снижение секреции |
—«— |
—«— |
|
потовые |
секреция холинергическая |
|
|
|
Метаболизм: |
|||
|
печень |
гликогенолиз, глюконеогенез |
|
|
|
жировые клетки |
липолиз |
β |
— |
|
секреция инсулина |
снижение |
β |
— |
и адреналин возбуждают только β—адренорецепторы, что
ведет к усилению сердечных сокращений и расширению бронхов. Учитывая, что
норадреналин вызывает наиболее сильное возбуждение β—адренорецепторов
миокарда и менее значительную реакцию (β—адренорецепторов гладких мышц
бронхов, трахеи, сосудов, первые называют адренорецепторами β1,
вторые — адренорецепторами β2.
Норадреналин, как и ацетилхолин,
способен не только вызывать эффект на постсинаптической мембране, но и одновременно
оказывать влияние на сам процесс выделения медиатора из пресинаптических
терминалей. Это достигается благодаря наличию в пресинаптических окончаниях тех
же, что и в постсинаптической мембране, адренорецепторов.
При взаимодействий норадреналина с
пресинаптическим α—адренорецептором выделение медиатора снижается.
Напротив, соединение норадреналина с пресинаптическим β—рецептором влечет
за собой усиление выделения медиатора. Вероятно, в обычных условиях такой
механизм работает по принципу обратной связи. Однако при сильном возбуждении
симпатического нейрона происходит чрезмерное повышение концентрации медиатора в
синаптической щели. В этом случае он взаимодействует с пресинаптическими а—адренорецепторами,
результатом чего является торможение выделения норадреналина (отрицательная
обратная связь). При слабом возбуждении преганглионарного нейрона выброс
норадреналина понижен и его концентрация в синаптической щели невелика. Это
влечет за собой возбуждение β—адренорецепторов и, как следствие, повышение
секреции медиатора (положительная обратная связь).
Саморегуляция выделения медиатора
касается не только уровня его содержания в синаптической щели. В мышцах
сосудов, бронхов, сердца известно также реципрокное торможение выделения
медиаторов из адренергических и холинергических окончаний. Так, при
одновременном возбуждении сердечных симпатических и парасимпатических волокон
наблюдается снижение выделения ацетилхолина. Это происходит в результате
возбуждения α—адренорецепторов пресинаптической мембраны холинергических
окончаний.
Наряду с этим возбуждение
сердечного холинергического нейрона вызывает, в свою очередь, в
постганглионарном симпатическом нейроне уменьшение выделения норадреналина.
Эффект опосредуется мускариновыми холинорецепторами пресинаптического
адренергического окончания. Эти наблюдения над тормозными отношениями
пресинаптических адренергических и холинергических окончаний указывают на то,
что взаимодействие между симпатической и парасимпатической нервной системой
осуществляется уже на уровне пресинаптических терминалей.
Норадреналин и адреналин в своем
действии на мембрану гладкомышечных клеток активируют находящуюся в ней аденилатциклазу.
Аденилатциклаза, в свою очередь, при наличии ионов магния катализирует
образование цАМФ (циклического 3', 5'—аденозинмонофосфата) из АТФ. Последний
активирует энергетический обмен, усиливая тем самым сердечную деятельность.
Вводимый извне норадреналин
вызывает повышение систолического и диастолического давления без ускорения
сердечного ритма, повышает силу сердечных сокращений. Вследствие сужения
почечных сосудов норадреналин вызывает торможение диуреза и задержку натрия
(см. разд. 12.9). Под влиянием норадреналина снижается секреция желудка и
кишки, расслабляется гладкая мускулатура кишки, в то время как слюноотделение
усиливается.
Адреналин
характеризуется более разнообразным диапазоном симпатического действия, чем
норадреналин. Благодаря одновременной стимуляции инотропной, хронотропной и
дромотропной функций (см. разд. 8.4.4) он повышает сердечный дебит. Действуя
непосредственно на бронхиальную мускулатуру, адреналин оказывает
бронхорасширяющее и антиспазматическое действие, рефлекторно снижает частоту и
амплитуду дыхания. Адреналин выступает и как антидиуретик, снижая выделение с
мочой ионов К+ и Na+ Он тормозит
желудочно—кишечную моторику, расслабляет стенки органов, но сокращает области
сфинктеров мочеполовой и кишечной систем, тормозит пищеварительную секрецию.
Адреналин повышает сократимость скелетных мышц, что особенно отчетливо
проявляется на фоне их утомления, т. е. оказывает действие, аналогичное
известному феномену Орбели—Гинецинского.
Для выполнения своих функций и
поддержания гомеостаза автономная нервная система наряду с обычными нейронами
содержит особые клетки, прием информации в которых осуществляется обычным
путем, а ответы —эндокринным способом. Такие клетки называются трансдукторами (от лат. transductio — перенос, перемещение).
К трансдукторам относят: 1)
хромаффинные клетки мозгового слоя надпочечников, которые отвечают на
холинергический передатчик преганглионарного симпатического волокна выделением
адреналина и норадреналина; 2) юкстагломерулярные клетки почки, которые
отвечают на адренергический передатчик постганглионарного симпатического
волокна выделением в кровяное русло ренина; 3) нейроны супраоптического и
паравентрикулярного ядер гипоталамуса, реагирующие на адренергический,
холинергический и другие передатчики выделением вазопрессина и окситоцина; 4)
нейроны ядер гипоталамуса, выделяющие в сосудистую систему факторы регуляции.
Трансдукторы содержат в соме и
аксоне хорошо идентифицируемые гистологически нейросекреторные тельца,
которые транспортируются по направлению от тела клетки. Аксоны этих клеток не
образуют синаптических контактов с другими нейронами или эффекторными органами.
Они свободно заканчиваются вокруг сосудов, с которыми образуют так называемые гемальные
органы.
Мозговое вещество надпочечников.
Центральную часть надпочечников составляет мозговое вещество, образованное из адреналовой
ткани. Клетки этого вещества содержат включения, окрашивающиеся в желто—коричневый
цвет двухромовокислым калием, что послужило поводом назвать их хромаффинными
клетками. Эти клетки расположены не только в мозговом слое надпочечников. В
виде скоплений они встречаются на поверхности аорты, в области каротидного
синуса, среди клеток симпатических ганглиев. В процессе эмбриогенеза
хромаффинные клетки образуются из тех же эктодермальных клеток, что и
симпатическая нервная система, и являются, по существу, гомологами
постганглионарного симпатического нейрона.
На хромаффинных клетках
синаптически оканчиваются преганглионарные симпатические нейроны, образующие
возбуждающие холинергические синапсы. При активации преганглионарных волокон в
кровь выделяется смесь, содержащая около 80% адреналина и 20% норадреналина.
Напротив, перерезка этих волокон прекращает секрецию катехоламинов. Нервные
центры, ответственные за секреторный процесс мозгового слоя надпочечников,
располагаются в гипоталамической области (см. разд. 6.2).
|
|
|
Рис.
5.13 Показатели изменения функций
различных органов под влиянием катехоламинов мозгового слоя надпочечников |
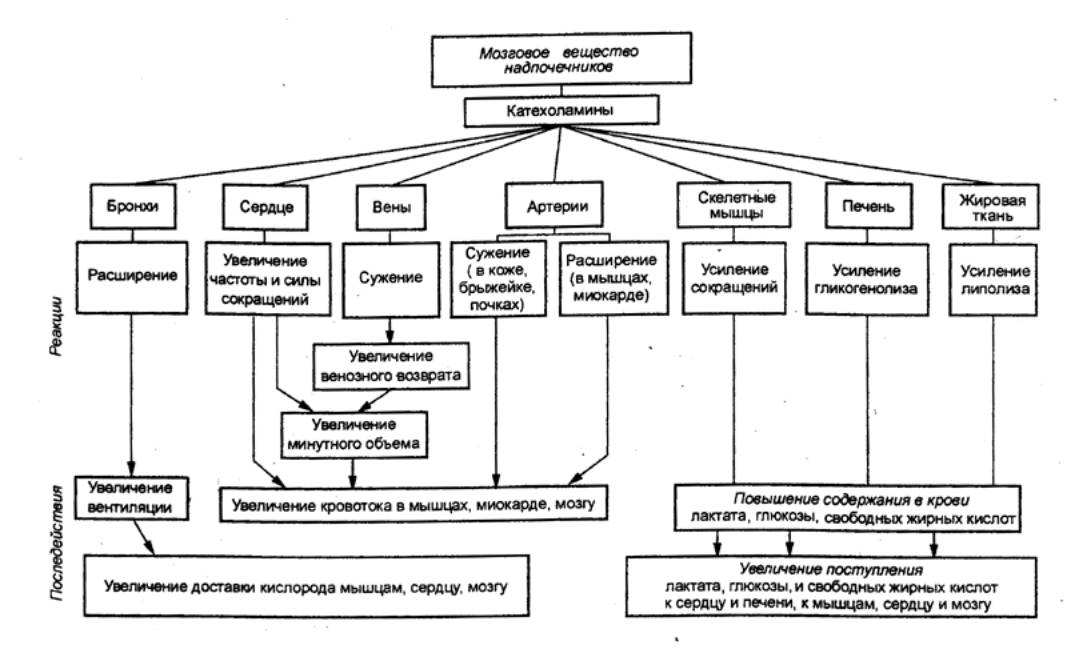
Точки приложения для продуцируемых
надпочечниками катехоламинов (рис. 5.13) те же, что и у симпатической нервной
системы, однако их выраженное действие проявляется главным образом в областях с
редкой адренергической иннервацией, таких как гладкомышечная стенка артериол, и
незначительно в областях с обильной иннервацией (семявыносящий проток). В
настоящее время катехоламинам мозгового слоя надпочечников отводят роль
метаболических гормонов, действие которых опосредуется в основном β—адренорецепторами.
Экстремальные ситуации типа
гипотермии, гипоксии, чрезмерного физического напряжения, а также
эмоционального состояния приводят к повышенному освобождению катехоламинов. Их
уровень, например, при эмоциональном стрессе может временно возрастать более
чем в 10 раз по сравнению с Уровнем покоя. Разумеется, при таком стрессорном
состоянии в реакцию вовлекается не только мозговое вещество надпочечников, но
прежде всего сама симпатическая нервная система и регулирующие ее деятельность
высшие центры, включая гипоталамус, лимбическую систему, кору больших полушарий
следовательно, в этой ситуации правильнее судить о реакции в целом всей симпатоадреналовой
системы.
В таких случаях
в результате прямого влияния симпатической нервной системы, а также выделения
катехоламинов мозговым слоем надпочечников создающим более продолжительное
действие, происходит увеличение транспорта кислорода и окисляемых
веществ к жизненно важным органам — мозгу сердцу. Взаимодействие катехоламинов
с β—адренорецепторами ведет к повышению содержания в крови глюкозы, лактата,
жирных кислот. Взаимодействие с α— и β—адренорецепторами приводит к
расширению артерий скелетных мышц и сердца, расширению бронхов, обширному
сужению вен, артерий кожи и внутренних органов (см. табл. 5.1).
В 1937 г. из энтерохромаффинных
клеток кишки и несколько позже из бычьей сыворотки был выделен энтерамин —
вещество, способное вызывать сокращение гладкой мускулатуры изолированных
органов. В химическом отношении он представляет производное индола — 5—гидрокситриптамин.
Название серотонин дано очищенному препарату.
Серотонин обнаруживают в тканях
практически всех видов животных, а также во многих растениях (в том числе
сливах, бананах, баклажанах). Общее количество серотонина в организме
млекопитающих достигает 10 мг. Из них почти 90% образуется в энтерохромаффинных
клетках слизистой оболочки пищеварительного тракта и откладывается в гранулы
цитоплазмы. Во время пищеварения часть вещества высвобождается в просвет
кишечника. Определенное количество его попадает в портальную систему (см. разд.
11.4.2).
В мозгу серотонин содержится
главным образом в структурах, имеющих отношение к регуляции висцеральных
органов. Особенно много его в лимбической системе, ядрах шва. Мозговой
серотонин, по—видимому, центрального происхождения. Он почти не проникает через
гематоэнцефалический барьер (см. разд. 3.27). Именно в нейронах богатых
серотонином областей обнаружены и ферменты, участвующие в его синтезе. Аксоны
этих нейронов проходят в бульбоспинальных путях и оканчиваются в сегментах
спинного мозга. Здесь они контактируют с клетками преганглионарных
симпатических нейронов и со вставочными нейронами студенистого вещества.
Высокая биологическая активность
серотонина определяет, вероятно, его незначительное содержание в тканях. Оно не
является даже приблизительно постоянным и зависит от вида, пола, возраста и
типа питания. Медиаторную функцию серотонина считают доказанной не только у
низших позвоночных, но и у млекопитающих.
На сосудистый аппарат
серотонин оказывает прямое и рефлекторное действие, что выражается в виде
вазоконстрикции или вазодилатации. Вазоконстрикция более отчетливо проявляется
на денервированных сосудах. В скелетных мышцах и кожных покровах преобладает
сосудорасширяющее действие, повышается капиллярная проницаемость. При прямом
действии серотонина возрастает сила сердечных сокращений, хотя этот эффект
маскируется баро— и хеморецепторными влияниями.
На дыхательный аппарат
вещество также оказывает прямое и рефлекторное действие. При прямом происходит сокращение
бронхиальной мускулатуры, сужение бронхов; при рефлекторном (в результате
стимуляции рефлексогенных зон и афферентных путей) — изменение частоты дыхания
и легочной вентиляции.
Неэффективные для сердечно—сосудистой
системы количества препарата отчетливо влияют на мускулатуру пищеварительного
тракта. Особенно чувствителен к серотонину человек: введение серотонина
вызывает у него начальную спастическую реакцию, переходящую в ритмические
сокращения с повышенным тонусом кишечной мускулатуры и завершающуюся
торможением спонтанной моторной деятельности. Являясь медиатором
метасимпатической нервной системы, серотонин выполняет также медиаторную
функцию в центральных образованиях.
Фармакологически установлено
существование трех типов серотонинергических рецепторов: D, М, Т. D—серотонинергические рецепторы
блокируются диэтиламидом лизергиновой кислоты. Они локализуются в основном в
гладкой мышце. Взаимодействие серотонина с этими рецепторами сопровождается сокращением гладких мышц. М—серотонинергические
рецепторы блокируются морфином. Они расположены в основном в ганглиях
автономной нервной системы. Влияя на эти рецепторы, серотонин вызывает
ганглиостимулирующий эффект. Т—серотонинергические рецепторы блокируются
типиндолом. Они обнаружены в сердечной и легочной рефлексогенных зонах.
Действуя на них, серотонин вызывает коронарный и легочный хеморефлексы.
Как и для других биологически активных веществ и медиаторов,
в автономной нервной системе обнаружено существование пресинаптических
серотонинергических рецепторов. Оказалось, что постсинаптические и
пресинаптические виды рецепторов, существование которых первоначально
установлено в автономной нервной системе, имеются также и в ЦНС на
соответствующих мембранах.
5.2.5. Аденозинтрифосфат (АТФ)
Роль АТФ в энергетическом обмене хорошо известна. Однако,
кроме того, он является синоптическим передатчиком, широко
представленным в различных органах и особенно в эффекторных нейронах
метасимпатической нервной системы, где локализуется в пресинаптических
терминалях. В связи с тем что при стимуляции этих терминалей выделяются
пуриновые продукты распада — аденозин и инозин, эта передача
получила название пуринергической.
Считают, что на ранних этапах эволюции АТФ был единственным
и общим для всех организмов медиатором. Впоследствии, с усложнением клеточной
организации, стали появляться ферментативные системы, синтезирующие новые
биологически активные соединения. Последние стали выполнять более узкие
медиаторные функции, однако АТФ их также сохранил.
Преобладающим действием АТФ на гладкой мышце является релаксация (от лат. relaxatio — ослабление напряжения), хотя иногда имеет место и возбуждающее действие. Оно
может быть прямое и непрямое. Прямое действие способны подавлять некоторые
препараты типа хинидина.
Исследования пуринергической передачи на лентах ободочной
кишки, мышцах желудка, тонкой и толстой кишки, пищевода, сфинктеров показали,
что раздражение пуринергического волокна сопровождается возникновением ТПСП. Ни
адреноблокаторы, ни десимпатизация не оказывают на этот ТПСП никакого влияния,
однако он полностью блокируется посредством тетродотоксина.
Пуринергические нейроны являются, по—видимому, главной
антагонистической тормозной системой по отношению к холинергической
возбуждающей системе, например, в механизме кишечной пропульсии (от лат. propello, propulsum — толкать вперед). Они участвуют в «нисходящем» торможении. Сокращения кишки,
следующие за возникающим в результате активации пуринергических структур
расслаблением, обеспечивают соответствующий механизм для прохождения пищевого
комка (болюса) по кишке. Пуринергические нейроны участвуют также в механизме
рецептивной релаксации желудка, расслаблении пищевого и анального сфинктеров.
Реакции гладкомышечных органов на АТФ опосредованы его
взаимодействием с пуринорецепторами клеточных мембран. На основании
фармакологических критериев выделено два их типа. Пуринорецепторы первого типа
более чувствительны к продукту распада АТФ — аденозину, второго — к самому АТФ.
Возможно, что роль рецептора пуринергической передачи выполняет
аденилатциклаза.
5.2.6.
Вероятные кандидаты в медиаторы
В числе возможных кандидатов в
медиаторы рассматривали большое количество биологически активных веществ:
глицин, ГАМК, субстанция Р, гистамин.
Распределение глицина в
спинном мозгу соответствует расположению тормозных интернейронов.
Ионофоретическое воздействие глицина на крестцовые парасимпатические нейроны
оказывает мощное угнетающее действие. Глициновая депрессия обратимо блокируется
стрихнином.
Тормозные эффекты на передачу возбуждения
в синапсах автономной нервной системы оказывает ГАМК. Считают, что она
может принимать участие в возникновении пресинаптического торможения.
Субстанцией Р наиболее
богаты стенки кишки, гипоталамическая область, особенно дорсальные корешки
спинного мозга. Последнее послужило одним из оснований к предположению, что она
может явиться медиатором чувствительных нервных клеток в области их
переключения на вставочные нейроны.
Во многих тканях организма
присутствует гистамин. Наибольшие его концентрации определяют в
желудочно—кишечном тракте, легких, кожных покровах. В нервной системе богатыми
гистамином областями являются постганглионарные симпатические волокна,
концентрация гистамина в которых достигает 100 мкг/г. В свободном состоянии
гистамин чрезвычайно активен и может вызывать разнообразные эффекты — снижение
кровяного давления, замедление сердечных сокращений, стимуляцию симпатических
центров.
Классическим действием гистамина
является повышение капиллярной проницаемости; кроме того, он вызывает сокращение
гладкой мускулатуры. На основании ответных реакций различных органов и тканей
на действие гистамина введено понятие об H1 и Н2—гистаминорецепторах.
Наряду с основной медиацией,
благодаря которой осуществляется передача влияний на эффекторы дуги автономного
рефлекса, существует большое число биологически активных веществ, получивших
название активных факторов, или местных гормонов. Это
простагландины, плазмакинины и др.; они оказывают значительное влияние на тонус
и действие автономной нервной системы.
Название простагландины (от лат. prostata — предстательная железа и glandula — железа) связано с
предстательными железами. Впервые простагландины были открыты в семенной жидкости
млекопитающих. В настоящее время различают большое число их разновидностей,
разделяемых на Е—, G—, А;
В—группы. Простагландины содержатся во всех тканях и органах, но наиболее
богата ими семенная жидкость. Основным местом их локализации в клетках является
жировой слой мембраны.
В структурах автономной нервной
системы, особенно в блуждающем нерве, содержится измеряемое количество
простагландинов. Они могут выделяться спонтанно или под влиянием стимуляции.
Большинство простагландинов оказывает возбуждающее действие на гладкую мышцу
(группа Е), угнетает желудочную секрецию (группы Е, А),
расслабляет бронхиальную мускулатуру (группа Е). На сердечно—сосудистую
систему простагландины оказывают разнонаправленное действие, зависящее от вида
животных: группы Е и А вызывают отчетливую вазодилатацию и
гипотензию, группа G таким
действием не обладает, вызывая вазоконстрикцию и сильную гипертензию.
Простагландины наделены отчетливым действием на почки и изменяют почечную
фильтрацию (А, Е). Им также отводят существенную роль в регуляции артериального
давления, которую они осуществляют совместно с ренин—ангиотензиновой системой
(см. разд. 6.4.2).
На основании вызываемых простагландинами эффектов
сформулировано представление о существовании простагландиновых рецепторов.
Попытки наделить простагландины медиаторной функцией не нашли
экспериментального подтверждения. Более вероятной кажется их роль в координации
нейрогуморальной передачи, в механизмах выделения и действия медиаторов.
Плазмакинины являются полипептидами, образующимися из плазматических
глобулинов. Эти вещества содержатся в тканях различных органов. Наиболее
известны три биологически активных плазмакинина: брадикинин, калидин и метионилкалидин.
Их вазодилататорный эффект почти в 10 раз превосходит действие гистамина и в
равной мере распространяется на сосуды скелетных мышц и внутренних органов, в
том числе на коронарные сосуды.
Даже в незначительных концентрациях плазмакинины вызывают
сокращение подвздошной кишки, матки, стимулируют кишечную моторику, повышают
тонус бронхиальной мускулатуры, увеличивают секрецию пищеварительных желез.
Посредством увеличения почечного кровотока и выделения ионов Na+, K+, а также воды они вызывают прямой
диуретический эффект.
К числу активных факторов относится и ренин—ангиотензиновая
система (см. разд. 6). Ренин является веществом прессорного
действия. Главное место его секреции — юкстагломерулярный аппарат почечных
клубочков. В комплексе ренин—ангиотензин ренин выступает как фермент, ангиотензин
наделен физиологическими свойствами. Наиболее выражены из них
кардиостимулирующее и вазоконстрикторное действия, превышающие по своей силе
влияние норадреналина более чем в 50 раз. Это действие может быть прямым и
опосредованным через симпатоадреналовую систему, которая способна
активироваться в этом случае непосредственно или центральными механизмами.
Наиболее сильно сосудосуживающее действие ангиотензина
проявляется во внутренних органах и кожных покровах, менее чувствительны к нему
сосуды скелетных мышц, мозга, сердца; почти не реагируют сосуды легких.
Особенно значительно проявляется его действие на почку, что выражается в
снижении почечной гемодинамики, нарушении клубочковой фильтрации и косвенном
влиянии альдостерона как регулятора канальцевой фильтрации и водно—электролитного
равновесия. Отмечены выраженные ганглиостимулирующие эффекты. Клеточное
действие ангиотензина сводится к активации АМФ и трансмембранного транспорта.
5.3. РЕФЛЕКТОРНЫЕ ПРОЦЕССЫ,
ВОЗНИКАЮЩИЕ ПРИ РАЗДРАЖЕНИИ ЧУВСТВИТЕЛЬНОГО ЗВЕНА АВТОНОМНОЙ ДУГИ
Процессы в автономной и соматической системах тесно связаны,
хотя при раздражении автономных чувствительных волокон обе системы вовлекаются
в ответ в разной степени. Рефлексы в этом случае разделяются на висцеро—висцеральные,
висцеросоматические и висцеросенсорные.
5.3.1. Висцеро—висцеральный рефлекс
Висцеро—висцеральный рефлекс включает
пути, в которых возбуждение возникает и заканчивается во внутренних органах. В
этом случае эффектор способен отвечать либо усилением, либо торможением
функций. Основой для осуществления этих процессов являются местные рефлекторные
дуги, замыкающиеся в узлах автономной нервной системы. Эти дуги могут быть
разного уровня:
одни из них замыкаются в
интрамуральных ганглиях, т. е. обеспечиваются метасимпатической нервной
системой, другие — в превертебральных ганглиях, третьи — в структурах более
высокого уровня.
Внутриклеточная техника
регистрации нейрональной активности показала, что, например, ободочный нерв
каудального брыжеечного симпатического ганглия содержит волокна, синаптически
контактирующие с постганглионарными клетками узла. Тем самым было подтверждено
существование синаптической связи в этом узле, обеспечивающей замыкание местной
рефлекторной дуги, рецептивным полем и эффектором которой является стенка
ободочной кишки.
Примером висцеро—висцерального
рефлекса может явиться классический опыт Гольца, показывающий, что
механическое раздражение брыжейки вызывает замедление сердечных сокращений;
ослабление тонуса мышц, суживающих зрачок при раздражении рецепторов желудочно—кишечного
тракта; изменение интенсивности дыхания, уровня кровяного давления, частоты
сердечных сокращений при раздражении каротидной и аортальной рефлексогенных
зон.
Дж. Ленгли, обнаружив установленное
Н. М. Соковниным (1877) явление истинного местного рефлекса, осуществляемого в
каудальном брыжеечном ганглии без участия ЦНС и представляющего собой передачу
возбуждения с чувствительного волокна на двигательную клетку, т. е. явление, не
укладывающееся в развиваемое им представление, дал ему иное толкование, назвав
этот вид аксон—рефлексом. С тех пор под аксон—рефлексом понимают
рефлекторные процессы, осуществляющиеся по разветвлениям аксона без участия
тела нейрона. В этом случае возбуждение якобы идет по одной ветви аксона, затем
переходит на другую ветвь и по ней эфферентно направляется к эффекторному
органу.
При раздражении волокон от кожных
болевых рецепторов возникает расширение сосудов и покраснение области,
иннервируемой этими волокнами. Тот же эффект сохраняется после удаления
спинного мозга. Его удается получить и после полной дегенерации симпатических
волокон, иннервирующих сосудистое русло исследуемого участка кожи. Эти
наблюдения послужили основанием считать, что в основе сосудистой реакции лежит
аксон—рефлекс. Такое объяснение хотя просто и понятно, но основано на косвенных
данных. Обеспечивающие аксон—рефлекс соответствующие структуры гистологически
пока не обнаружены, и само явление не подвергнуто детальному
нейрофизиологическому анализу.
В ганглиях автономной нервной
системы аксон—рефлекс возникает только в искусственной обстановке при
электрическом раздражении. Поэтому для того чтобы установить, является ли он
истинным, присущим этому отделу нервной системы явлением, необходимо
воспроизведение этого рефлекса в адекватных условиях при действии раздражителя
на адекватно воспринимающую поверхность.
Существует и иное объяснение
эффектов, объединяемых понятием аксон—рефлекса. Считают, что при возбуждении
кожных болевых рецепторов из их мембран происходит выделение биологически
активных веществ типа субстанции Р или АТФ, обладающих сосудорасширяющим
действием. Основанием к такому заключению является то, что эти и подобные им
вещества действительно обнаружены в рецепторах и чувствительных волокнах. Кроме
того, при возбуждении
волокон и рецепторов концентрация в них указанных биологически активных веществ
резко снижалась.
В литературе, касающейся функциональной структуры
висцеральной периферии, часто встречаются необычные терминологические
конструкции типа «эфферентная функция чувствительных нервных волокон». Несмотря
на кажущуюся абсурдность, за этим термином при внимательном рассмотрении
отчетливо вырисовывается новый класс реальных явлений, не укладывающихся в
классические понятия «чувствительная иннервация», «двигательная иннервация»
висцеральных органов.
Возникает вопрос, не означает ли это еще один шаг к размытию
понятия «классический нейроэффекторный синапс» или это открытие новых
функциональных возможностей интероцепторов, а стало быть природы и механизмов
аксон—рефлекса. Последнее имеет большее право на существование, так как
основным субстратом, своеобразным местом действия вновь установленных эффектов
является интероцептор, обладающий, на первый взгляд, несколько неожиданным
свойством. Свойство это заключатся в том, что в ответ на адекватную стимуляцию
или раздражение электрическим полем рецептор высвобождает определенные
химические вещества, оказывающие специфическое действие на окружающую ткань.
Такими веществами являются преимущественно нейропептиды. Эти
закономерности относятся не только к чувствительным терминалям специфической
популяции первичных сенсорных нейронов, тела которых расположены в спинальных
ганглиях, но и к сенсорным окончаниям функциональных модулей метасимпатической
нервной системы.
Опираясь на существующие
экспериментальные данные, общую схему функционирования этих нейронов можно
представить следующим образом. Синтезирующиеся в соме сенсорного нейрона
неиропептиды благодаря аксонному транспорту перемещаются в его терминали. Из
них наиболее часто выявляемым пептидом является субстанция Р. Для того
чтобы представить механизм использования нейропептидов, предполагается, что в
этом случае происходит возбуждение интероцептивной терминали, например, в ответ
на растяжение стенки органа, приводящее к такой последовательности возникающих
событий.
Во—первых, возбуждение
интероцептора само по себе является стимулом к локальному выделению
нейропептидов из их тактического пула, находящегося в его терминальных
участках. Во—вторых, при наличии коллатерали по ходу сенсорного волокна и при
достаточно высоком факторе надежности проведения в месте бифуркации возбуждение
может перейти, на эту коллатераль, антидромно достичь другого терминального
участка и вызвать такое же локальное, но уже пространственно удаленное
от места раздражения выделение нейропептидов. Именно таким образом возникает
аксон—рефлекс в его классическом понимании. В—третьих, дойдя до образований
ганглионарного или спинального уровня, формирующих аксодендритные или
аксосоматические контакты на интернейронах дорсальных рогов, возбуждение также
вызывает выделение нейропептидов. Однако непосредственной мишенью в этом случае
является уже нейрон второго уровня (проекционный нейрон) и передача возбуждения
идет в рамках структуры обычного классического межнейронного синапса. И в
первом, и во втором случаях существует целая группа клеток—мишеней, к которым
выделившиеся неиропептиды диффундируют через межклеточное пространство,
окружающее гладкомышечные, а также эпителиальные клетки стенок полых
висцеральных органов и сосудистого дерева клетки местных эндокринных
образований и т. д.
5.3.3. Висцеросоматический рефлекс
Висцеросоматический рефлекс включает пути, по которым
возбуждение в дополнение к висцеральным вызывает также соматические ответы в
виде, например, усиления (сокращения) или торможения текущей активности
скелетных мышц. Известно, что раздражение хемо— и механорецёпторов каротидной
зоны сопровождается торможением общей двигательной активности организма, а
интенсивное раздражение рецепторов пищеварительного тракта может вызвать
сокращение мышц брюшного пресса или движение конечностей.
В силу сегментарной организации автономной и соматической
иннервации при заболеваниях внутренних органов в ограниченных участках кожи
возникает повышение тактильной и болевой чувствительности (рис. 5.14). Эти боли
называют отраженными, а области, в которых они появляются, — зонами
Захарьина—Геда. В рассматриваемом случае висцеральные и кожные
чувствительные волокна конвергируют на одних и тех же нейронах спинно—таламического
пути.
|
|
|
Рис. 5.14 Пути возникновения отраженной боли. Слева
изображены некоторые из ноцицептивных афферентов внутренних органов,
оканчивающихся в заднем роге на тех же нейронах, что в ноципептивные
афференты кожи; справа — в некоторых случаях ветви одного и того же
ноципептивного афферента иннервируют и поверхностную, и глубокую ткани. |
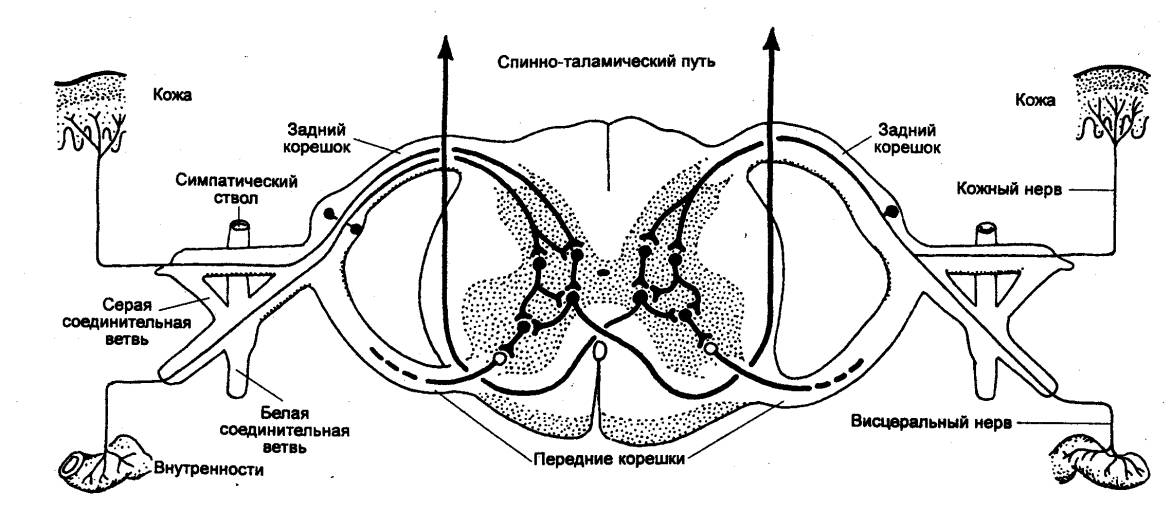
Во влиянии с рецептивных полей внутренних органов на скелетно—мышечную
систему принято различать (В. Н. Черниговский) корригирующие и пусковые
влияния. Первые приводят к изменению сокращений скелетных мышц, уже
происходящих под влиянием других сенсорных стимулов, вызывая либо усиление,
либо угнетение. Вторые самостоятельно вызывают активность скелетно—мышечной
системы. Оба вида влияния связаны с поступлением сигналов по чувствительным
путям автономной рефлекторной дуги. Нередко пусковые влияния дополняются
корригирующими.
5.3.4. Висцеросенсорный
рефлекс
Висцеросенсорный рефлекс включает
пути, в которых в ответ на раздражение автономных чувствительных волокон
возникают реакции не только во внутренних органах, мышечной системе, но и
изменяется соматическая чувствительность. В этом случае зона повышенного восприятия,
как правило,
|
|
|
Рис. 5.15 Афференты и структуры соматовисцеральной системы в стволе головного
мозга (справа схема организации — вид сзади) Афференты
тройничного нерва и трех его ветвей (V1, V2, V3). блуждающего и
языкоглоточного нервов, центральная афферентная система тройничного нерва и
ее восходящие пути, ретикулярная формация и пути, приходящие из спинного
мозга. |
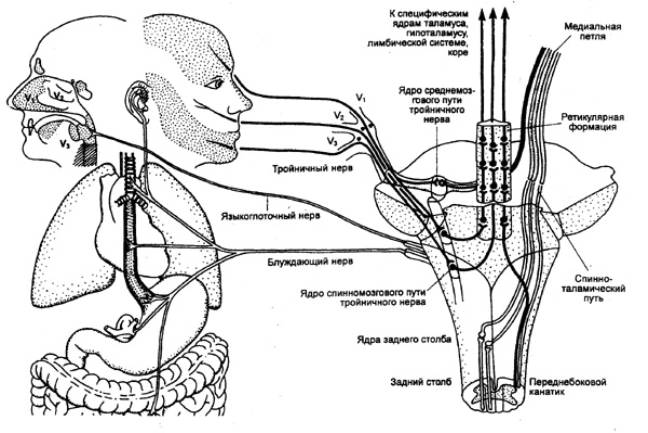
ограничивается
участком кожи, иннервируемым сегментом, к которому поступают импульсы от раздражаемого
внутреннего органа. Для вызова таких рефлексов необходимо чрезвычайно сильное и
продолжительное раздражение.
Существует и соматовисцеральный рефлекс. Он включает
пути (рис. 5.15), вовлекающие активацию экстероцепторов и соматических
афферентных волокон и заканчивающиеся в висцеральных органах, вызывая изменение
их функций. Примером может служить сосудистая реакция при термических
воздействиях на рецепторы кожи.
5.4.
ВЛИЯНИЕ АВТОНОМНОЙ НЕРВНОЙ СИСТЕМЫ НА ДЕЯТЕЛЬНОСТЬ ЭФФЕКТОРНЫХ ОРГАНОВ
Автономная нервная система
координирует и адаптирует деятельность органов, участвующих в сохранении
динамического равновесия жизненных функций, регулируя метаболизм, возбудимость,
автоматизм внутренних органов и самой ЦНС. Многие внутренние органы имеют
двойную и даже тройную — симпатическую, парасимпатическую и метасимпатическую —
иннервацию. Изучение роли каждой из них обычно осуществляют посредством
электрического раздражения, хирургического или фармакологического выключения.
Раздражение симпатических
волокон вызывает учащение работы сердца, увеличение его ударного объема,
расслабление бронхиальной мускулатуры, снижение перистальтики желудка и кишки,
расслабление жёлчного пузыря, сокращение сфинктеров и т. д. Стимуляция парасимпатического блуждающего нерва сопровождается
противоположным эффектом: уменьшается частота сокращений сердца, ослабевает
сила сокращения предсердий, сокращается бронхиальная мускулатура, усиливается
моторика желудка и кишки, сокращается жёлчный пузырь, расслабляются сфинктеры
(см. табл. 5.1).
В физиологических условиях между
тонусом этих двух отделов (см. ниже) имеет место «относительное равновесие».
При преобладании тонуса одного из них тонус другого снижается.
Аналогия с известным для скелетной
мышечной системы принципом реципрокности послужила основанием для представления
об «антагонистических» отношениях между симпатической и парасимпатической
системами. При этом взаимодействия между ними уподоблялись коромыслу весов, в
которых подъем на определенный уровень одной чаши влечет за собой снижение
точно на такую же величину другой. Считали даже, что функция органов может
протекать нормально лишь при уравновешенности одних влияний другими.
Однако такому представлению
противоречит ряд фактов. Например, слюноотделение стимулируется и симпатическими,
и парасимпатическими волокнами: раздражение парасимпатического нерва —
барабанной струны —вызывает обильное выделение жидкой слюны; при раздражении
симпатического нерва слюна тоже образуется, но в небольших количествах и со
значительным содержанием органических веществ. Таким образом, здесь проявляется
согласованная реакция, необходимая для пищеварения. Кроме того, существует и
ряд органов, снабжаемых только симпатическими или только парасимпатическими
волокнами. Это многие кровеносные сосуды, селезенка, гладкие мышцы глаза,
волосяных луковиц, некоторые экзокринные железы (см. табл. 5.1).
Отделы симпатической и
парасимпатической нервной системы в большом числе случаев оказываются
функционально синергичными. Наиболее отчетливо это прослеживается при рассмотрении
рефлексов с барорецепторов на сердце (см. разд. 8.4.9). Возникающее в
результате повышения артериального давления возбуждение барорецепторов вызывает
снижение силы и частоты сокращений сердца. Наблюдаемый эффект связан с
возрастанием активности вагусных (блуждающего нерва) сердечных волокон, а также
со снижением активности симпатических волокон.
5.4.1.
Адаптационно—трофическая функция симпатической нервной системы
Предложенная Дж. Ленгли
классическая схема распространения симпатической иннервации предусматривала ее
влияние только на гладкую мускулатуру и железы. Однако симпатические импульсы
могут оказывать влияние и на скелетные мышцы. Если стимуляцией двигательного
нерва довести мышцу лягушки до утомления (рис. 5.16), а затем одновременно раздражать
симпатический ствол, то работоспособность утомленной мышцы повышается — феномен
Орбели—Гинецинского. Сама по себе стимуляция симпатических волокон не
вызывает сокращения мышцы, но изменяет состояние мышечной ткани, повышает ее
восприимчивость к передаваемым по соматическим волокнам импульсам. Такое
повышение работоспособности мышцы является результатом стимулирующего влияния
обменных процессов в мышце: растет потребление кислорода, увеличивается
содержание АТФ, креатинфосфата, гликогена. Полагают, что местом приложения
этого влияния является нервно—мышечный синапс.
Было также обнаружено, что
стимуляция симпатических волокон может значительно изменять возбудимость
рецепторов и даже функциональные свойства ЦНС. Например, при раздражении
симпатических волокон языка возрастает
|
|
|
Рис. 5.16 Феномен Орбели—Гинецинского
— усиление сокращения утомленной мышцы лягушки при раздражении симпатического
нервного ствола Отметка раздражения указана на верхней
линии, сокращения мышцы вызываются ритмическими раздражениями двигательных
нервных волокон. |

вкусовая
чувствительность, при раздражении симпатических нервов наблюдается повышение
рефлекторной возбудимости спинного мозга, изменяются функции продолговатого и
среднего мозга. Характерно, что при разной степени возбуждения симпатическая
нервная система оказывает на органы и ткани однотипные влияния. Удаление
краниальных шейных симпатических узлов у животных приводит к уменьшению
величины условных рефлексов, хаотичности их протекания, преобладанию в коре
больших полушарий процессов торможения.
Эти факты были обобщены Л. А. Орбели в теории адаптационно—трофической
функции симпатической нервной системы, согласно которой симпатические
влияния не сопровождаются непосредственно видимым действием, но значительно
изменяют функциональную реактивность или адаптивные свойства тканей.
Симпатическая нервная система активирует деятельность
нервной системы в целом, активирует защитные функции организма, такие как
иммунные процессы, барьерные механизмы, свертывание крови, процессы
терморегуляции. Ее возбуждение является непременным условием любых стрессорных
состояний, оно служит первым звеном запуска сложной цепи гормональных реакций.
Особенно ярко участие симпатической нервной системы
обнаруживается в формировании эмоциональных реакций человека независимо от
вызвавшей их причины. Так, радость сопровождается тахикардией, расширением
сосудов кожи, страх — замедлением сердечного ритма, сужением кожных сосудов,
потоотделением, изменением кишечной перистальтики, гнев — расширением зрачков.
Следовательно, в процессе эволюционного развития
симпатическая нервная система превратилась в особый инструмент мобилизации всех
ресурсов (интеллектуальных, энергетических и др.) организма как целого в тех
случаях, когда возникает угроза самому существованию индивидуума.
Такое положение симпатической нервной системы в организме
опирается на обширную систему ее связей, позволяющую посредством мультипликации
импульсов в многочисленных пара— и превертебральных ганглиях мгновенно вызывать
генерализованные реакции практически всех органов и систем. Значительным
дополнением является и выброс в кровь из надпочечников и хромаффинной ткани
«жидкости симпатической нервной системы» — адреналина и норадреналина.
В проявлении своего возбуждающего действия симпатическая
нервная система приводит к изменению гомеостатических констант организма, что
выражается в повышении кровяного давления, выходе крови из кровяных депо,
поступлении в кровь ферментов, глюкозы, повышении метаболизма тканей, снижении
мочеобразования, угнетении функции пищеварительного тракта и т. д. Поддержание
постоянства этих показателей целиком ложится на парасимпатическую и
метасимпатическую части.
Следовательно, в сфере управления
симпатической нервной системы находятся в основном процессы, связанные с
расходом энергии в организме, парасимпатической и метасимпатической — с ее
кумуляцией.
Значение симпатической нервной
системы убедительно демонстрируется в опытах с ее хирургическим,
химическим или иммунным удалением. Полная экстирпация у кошек симпатических
стволов, т. е. тотальная симпатэктомия, не сопровождается значительными
расстройствами висцеральных функций. Артериальное давление находится почти в
нормальных пределах, исключая небольшую недостаточность, возникающую из—за
выключения рефлексогенных зон; в близких к нормальным пределах развертывается
функция пищеварительного канала, продолжают оставаться возможными
репродуктивные функции: оплодотворение, беременность, роды. И тем не менее
симпатэктомированные животные не в состоянии осуществлять физические усилия, с
большим трудом оправляются от кровотечений, расстройств аппетита, шока,
гипогликемии, а также плохо переносят охлаждение и перегревание. У
симпатэктомированных животных не бывает проявления характерных защитных реакций
и показателей агрессивности: тахикардии, расширения зрачков, повышения притока
крови к соматической мускулатуре.
Рядом преимуществ обладает иммуносимпатэктомия.
Не оказывая значительного влияния на физическое развитие и общеповеденческие
реакции животных, этот метод вместе с тем позволяет получить своеобразную
модель для изучения функции автономной нервной системы в хронических условиях.
Определенным преимуществом является то, что введение фактора роста нервов в
условиях атрофии симпатической нервной системы позволяет получить на одних и
тех же животных и ее гипертрофию, создавая, таким образом, редкий в
экспериментальных условиях двойной контроль.
После перерезки симпатических
волокон и их дегенерации иннервируемые органы могут в какой—то мере
атрофироваться. Однако спустя несколько недель после денервации возникает их
повышенная чувствительность к медиаторам и веществам медиаторного типа. Этот
эффект наглядно прослеживается на зрачке животного после удаления краниального
шейного симпатического ганглия. Обычно вслед за операцией в результате
преобладания парасимпатического тонуса происходит сужение зрачка. Спустя
определенное время его величина приближается к исходной, а в условиях
эмоционального напряжения даже резко увеличивается.
Этот факт объясняют возникновением
сенситизации (гиперчувствителъности) денервированной мышцы к адреналину
и норадреналину, выбрасываемым из надпочечников в кровь во время эмоций.
Вероятно, в основе этого явления лежит изменение способности мембран
денервированных клеток к связыванию кальция и изменению проводимости.
5.4.2.
Роль парасимпатической нервной системы в регуляции висцеральных функций
Области влияния парасимпатической
нервной системы на процессы в организме сравнительно ограничены (см. табл.
5.1). Эти влияния могут сказываться либо прямо на иннервируемых органах, как в
кольцевой мускулатуре радужной оболочки глаза или в слюнных железах, либо через
посредство метасимпатической нервной системы. В первом случае постганглионарный
нейрон сам непосредственно контактирует с эффектором и вызываемое, им действие
зависит главным образом от прямых влияний структур ЦНС.
Во внутренних органах
преганглионарное парасимпатическое волокно оканчивается не на эффекторе —
мышечных волокнах или железистых клетках, а на интернейроне или эфферентном
нейроне метасимпатической нервной системы, который представляет общий
конечный путь для импульсов, поступающих по блуждающему и тазовому нервам
(см. рис. 5.4). Здесь они вступают во взаимодействие с импульсами, посредством
которых осуществляются процессы базовой внутриорганной местной
метасимпатической регуляции.
Таким образом, парасимпатические
влияния оказываются не прямыми, а опосредованными. Поэтому результат
адекватного раздражения (в отличие от чрезмерной электрической стимуляции
одновременно всех вагусных волокон) не бывает однозначным. Он зависит от
текущих внутриорганных процессов. Тут могут возникать возбуждение или
торможение функции органа, включаться или выключаться различные регуляторные
влияния, направленные на поддержание нормальной деятельности, стабилизацию
гомеостатического состояния.
5.4.3.
Участие метасимпатической нервной системы в регуляции висцеральных функций
Давно известно, что многие внутренние
органы, извлеченные из организма, при создании соответствующих условий
продолжают осуществлять присущие им функции без особых видимых изменений,
например сохраняются перистальтическая и всасывательная функции у кишки,
сокращается перфузируемое сердце. Сегменты или полоски стенок мочеточника,
матки, жёлчного пузыря также продолжают сокращаться с частотой и амплитудой,
характерной для каждого органа. Эта относительная функциональная автономия
объясняется наличием в их стенках метасимпатической нервной системы,
обладающей собственным нейрогенным ритмом и имеющей необходимые для
самостоятельной рефлекторной деятельности звенья — сенсорное, ассоциативное,
эфферентное и медиаторное.
Метасимпатическую нервную систему
рассматривают сейчас как сугубо периферическую, независимую интегративную
систему, способную самостоятельно обрабатывать сенсорную информацию и
генерировать организованный выход, который контролирует и координирует
поведение эффекторов. Существующие данные свидетельствуют о том, что принципиальная
модель метасимпатической нервной системы полностью совпадает с подобной моделью
центральной нервной системы.
Действительно, в составе
метасимпатической нервной системы существуют собственные сенсорные элементы, которые
могут быть механо—, хемо—, термо— и осморецепторами. Они постоянно посылают в
свои внутренние сети информацию о состоянии стенки внутреннего органа. Наряду с
этим сенсорные элементы способны передавать сигналы и в центральные структуры
нервной системы.
Метасимпатические нейроны
синаптически связываются в сети, которые обрабатывают поступающую сенсорную
информацию и одновременно контролируют активность эффекторных нейронов.
Последние, в свою очередь, являются общими конечными путями к эффекторам и могут
инициировать, поддерживать, наконец, тормозить их поведение.
У позвоночных животных ядерные
образования симпатической и парасимпатической систем располагаются в ЦНС.
Метасимпатическая нервная система не имеет таких представительств и является в
значительно большей степени, чем обе названные, самостоятельной, даже если
автономию понимать не в относительном, а в преувеличенном, но не
абсолютизированном до крайности смысле. Симпатическая нервная система
иннервирует все сосуды, а через них
скелетные мышцы, все внутренние органы, часть гладкой
мускулатуры глазного яблока, мышцы, поднимающие волосы, и т. д. Область
иннервации парасимпатической нервной системы более узкая: экзокринные железы,
часть гладкой мускулатуры глазного яблока, внутренние органы. Сфера иннервации
метасимпатической нервной системы еще более ограниченна и охватывает сугубо
внутренние органы и то не все, а лишь обладающие собственным моторным ритмом.
Для этих органов метасимпатическая
иннервация является базовой. Все звенья ее рефлекторного пути
локализуются только в интрамуральных ганглиях. Она не имеет своего центрального
аппарата. В результате такой организации ее эфферентные связи с центральными
структурами опосредованы нейронами симпатической и парасимпатической нервной
системы, образующими синаптические контакты на телах и отростках
метасимпатичееких интернейронов и эффекторных нейронов.
Представление о том, что
метасимпатическая нервная система — это диффузный парасимпатический ганглий, в
котором прямые синаптические контакты между преганглионарными волокнами и
постганглионарными клетками являются основой для контроля, например,
желудочной, кишечной или сердечной функции, при экспериментальном рассмотрении
оказалось несостоятельным. Так же несостоятельно и представление о
метасимпатической нервной системе как третьем нейроне в эфферентном звене
симпатического пути.
Метасимпатическая нервная система —
это относительно независимая самостоятельная интегративная система.
Такая относительная независимость
имеет ряд эволюционных преимуществ. Они заключаются в более эффективном
контроле и управлении обширными висцеральными системами и территориями,
исключающими, в частности, необходимость существования большого числа длинных
соединительных путей между ЦНС и висцеральными органами. Не менее существенным
является и уменьшение пространства в ЦНС, которое в противном случае было бы
занято обработкой излишней информации.
Очевидность невыгодности размещения
в ЦНС аппарата, необходимого для постоянного и непрерывного контроля за каждой
из висцеральных функций, подтверждается еще и тем, что, например, только одна
кишечная часть метасимпатической системы имеет то же число клеток (108),
что и весь спинной мозг, а количество метасимпатичееких нейронов, приходящихся
на 1 см2 поверхности кишки, составляют около 20 000.
Существование местных
метасимпатичееких механизмов регуляции функций висцеральных органов имеет
определенный физиологический смысл. Кроме освобождения ЦНС от избыточной
информации их наличие увеличивает надежность регуляции функций. Эта
регуляция без особых существенных изменений может осуществляться даже при
полном выключении связи с центральными структурами.
Прибегая к аналогии, функцию
метасимпатической нервной системы можно уподобить микропроцессору,
расположенному в непосредственной близости от эффекторов (гладкая мышца,
экскретирующий и всасывающий эпителии, эндокринные элементы, иммунные
структуры), которые ею контролируются и регулируются. Это местная система.
Именно она инициирует и программирует работу эффекторов, осуществляя текущую
регуляцию физиологических процессов и обеспечивая в полном объеме поддержание
постоянства внутренней среды организма. Что касается ЦНС, или «основного
компьютера», то он также получает информацию от висцеральных органов,
перерабатывает ее и передает команды нейронам метасимпатической сети.
Эти команды соответствуют состоянию отдельного органа или органов, а также
поведенческому состоянию индивидуума.
Нейробиологические свойства
метасимпатических сетей во многом напоминают организацию центральных нервных
структур позвоночных. По своей композиции, нейрохимии, иммунореактивности
протеинов, обмену материалами с циркуляторной системой, интегративной функции
они сходны с ядерными образованиями мозга млекопитающих и напоминают вынесенные
на периферию нервные центры.
Глиальные элементы
метасимпатической нервной системы, подобно астроглии головного мозга, в большом
количестве окружают каждый нейрон. Существенным моментом является наличие в тех
и других структурах плотного нейропиля, так как известно, что во всех системах
интеграции обработка значительной части информации происходит в микроцепях
именно синаптического нейропиля.
|
|
|
Рис. 5.17 Типы.
метасимпатических нейронов млекопитающих А —мотонейроны, идущие к гладкой
мышце; Б — интернейроны; В —
парасимпатические и симпатические входы; Г — иннервация местного кровотока. Медиаторы: АТФ —
аденозинтрифосфат, АХ — ацетилхолин, ВИП — вазоактивный интестинальный
пептид, НА — норадреналин. Сер — серотонин, СП — субстанция Р. Виды нейронов:
Адр — адренергический, НАНХ — неадренергический, нехолинергический, Сер —
серотонинергический, Хол —холинергический, знаками «+» и «—» показана
направленность действия. |
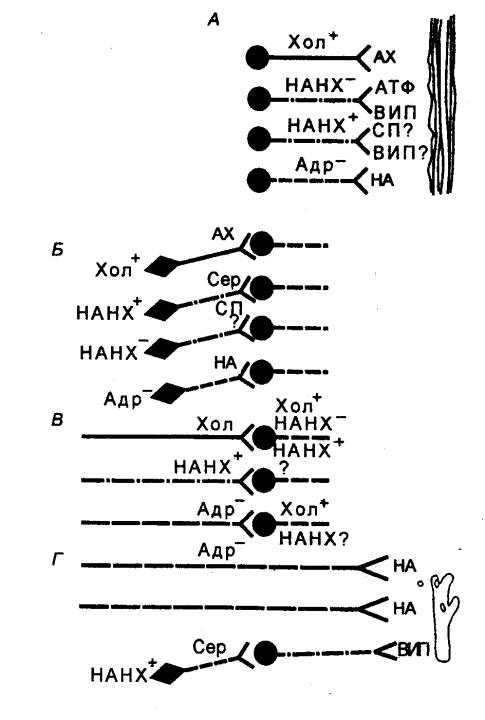
Большинство реальных и
предполагаемых медиаторов метасимпатической нервной системы является и
нейротрансмиттерами в ЦНС. Сейчас твердо установлено, что, например, в
энтеральной части метасимпатической нервной системы медиаторную функцию
осуществляет не менее 20 медиаторов и модуляторов холинергической,
адренергической, серотонинергической, пуринергической, пептидергической,
дофаминергической, ГАМК—ергической и другой природы (рис. 5.17). Все это —
основа для широкого диапазона многообразных регуляторных воздействий.
В метасимпатической нервной системе
содержатся также гематоганглионарные барьерные структуры, напоминающие по своей
функции гематоэнцефалический барьер. Они защищают метасимпатические нейроны от
непосредственного воздействия веществ, циркулирующих в крови.
По современным представлениям,
основная роль парасимпатической и метасимпатической систем состоит в
осуществлении механизмов различных функций, обеспечивающих гомеостаз —
относительное динамическое постоянство внутренней среды и устойчивость основных
физиологических функций.
В отличие от них симпатическая
нервная система рассматривается как система тревоги, мобилизации
защитных сил и ресурсов для активного взаимодействия с факторами среды. Из
этого следует, что, обеспечивая максимальное напряжение функций органов и
систем для защиты организма, симпатическая нервная система дестабилизирует
постоянство внутренней среды. Задачу восстановления и поддержания этого
постоянства, нарушенного в результате возбуждения симпатической нервной
системы, берут на себя парасимпатическая и метасимпатическая системы.
Многие симпатические и
парасимпатические эфферентные волокна, а также эфферентные клетки
метасимпатической нервной системы находятся в состоянии непрерывного
возбуждения, получившего название тонус. Для явлений, обозначаемых этим
термином, например, состояния нейронов, иннервирующих сосуды, сердце и другие
полые органы, обязательно длительное поддержание постоянного внешнего эффекта.
Тонус — одно из проявлений гомеостаза в организме и одновременно один из механизмов
его стабилизации.
Тонус покоя можно измерить либо
прямым методом посредством регистрации активности отдельных волокон и клеток,
либо косвенно — по показателям изменения деятельности исполнительного органа
после электрической стимуляции иннервирующих его волокон. Величина тонической
импульсной активности в пре— и постганглионарных волокнах находится в пределах
от 0,1 до 5,0 Гц и зависит от состояния иннервируемых гладких мышц, имеющих,
кроме того, собственный базальный мышечный тонус (см. разд. 1.2.8 и 8.4.6).
Тонические влияния симпатической
нервной системы, так же как и блуждающего нерва, наиболее подробно исследованы
на сердце. Преобладание симпатического тонуса обычно оценивают на основании
таких показателей, как учащенный сердечный ритм, незначительность его изменения
при атропинизации, слабая выраженность глазосердечного и синокаротидного
рефлексов. В сердечных симпатических волокнах поток непрерывных эфферентных
разрядов обычно снижается во время вдоха. Характерным является отчетливое
группирование импульсов синхронно систолическим сокращениям.
Симпатической нервной системе
принадлежит ведущая роль в создании общего сосудистого тонуса (рис.
5.18). В зависимости от тонуса сосудодвигательных центров и уровня кислых
катаболитов в тканевой жидкости симпатические волокна, иннервирующие мелкие
сосуды и сосуды среднего диаметра, приспосабливают кровоток к местным и общим
нуждам организма. В тех случаях, когда в системном давлении обнаруживается
тенденция к снижению, происходит включение компенсаторных адренергических
механизмов. Местные сосудосужающие реакции усиливаются одновременным выбросом
адреналина, возникающим в результате прихода к надпочечникам эфферентных
разрядов сосудодвигательного центра.
|
|
|
Рис.
5.18 Зависимость
гидродинамического сопротивления в сосудах скелетных мышц конечности кошки от частоты надпороговых раздражений преганглионарных волокон
поясничного отдела симпатического ствола. |
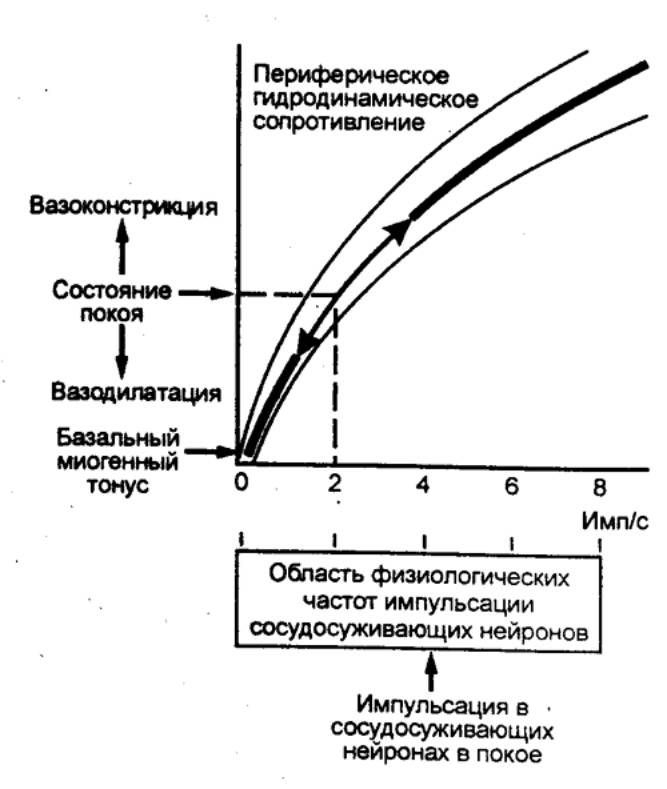
Иннервирующие сердце парасимпатические
волокна также находятся в состоянии непрерывного возбуждения, и передаваемая по
ним информация направлена на снижение частоты сердечных сокращений. Тоническая
импульсация в волокнах блуждающего нерва постоянно оказывает на сердце отрицательное
инотропное действие, снижая интенсивность сокращений желудочков. Напротив,
ослабление тонуса способствует учащению сердечных сокращений. Характерно, что
возникающая после перерезок блуждающих нервов тахикардия может наступать у
предварительно десимпатизированных животных. Следовательно, она не
обусловлена возрастанием симпатической активности, а полностью зависит от
интенсивности притока по волокнам блуждающих нервов тормозных влияний.
Роль тонуса блуждающих нервов состоит, по—видимому,
в создании в сердце значительных резервов, которые используются организмом в
ситуациях, сопровождающихся повышенной сердечной деятельностью. Именно снижение
этого тонуса, ведущее к возрастанию частоты сердечных сокращений, является
основой для быстрой перестройки режима работы органа. Вероятно, поэтому эффекты
ваготомии (пересечения блуждающего нерва) наиболее отчетливо проявляются у
животных, способных к быстрой смене мышечной деятельности (зайцы, обезьяны).
Тоническая неравнозначность симпатической
и парасимпатической частей автономной нервной системы явилась основанием для
создания своеобразной конституционной классификации, в которой
преобладание тонуса симпатической нервной системы обозначается как симпатикотония,
парасимпатической — ваготония. В этих случаях главным показателем
является уровень деятельности иннервируемых органов. Наиболее характерными
проявлениями у симпатикотоников являются учащенный пульс, отсутствие
потливости. У ваготоников наблюдается склонность к покраснениям, замедленному
пульсу, потливости, необъяснимым желудочным расстройствам. Чистые формы
симпатикотонии и ваготонии встречаются довольно редко.
О природе тонической активности в
симпатических и парасимпатических эфферентных волокнах известно мало. Она в
основном зависит от уровня импульсации, следующей в ЦНС от рецептивных полей
рефлексогенных зон висцеральных органов, а также соматических рецепторов.
Прямым свидетельством того является возникновение тахикардии при денервации
аорты и каротидного синуса, а также исчезновение разрядов в сердечных ветвях
блуждающих нервов при снижении артериального давления. По—видимому, в этом
процессе участвует и сигнализация от тканевых рецепторов.
Тонус симпатических и
парасимпатических эфферентных волокон связан, помимо того, еще и со спонтанной
деполяризацией нейронов—пейсмекеров, локализованных в основном в
структурах продолговатого мозга.
В метасимпатической нервной системе
основными элементами, задающими ритм и постоянно поддерживающими тонус
функционального модуля, являются специальные клетки—осцилляторы. Эти
нейроны характеризуются разрядами взрывного типа. На их функцию не оказывают
влияния блокаторы синаптической передачи и вещества медиаторного типа.
5.5. ЦЕНТРЫ
РЕГУЛЯЦИИ ВИСЦЕРАЛЬНЫХ ФУНКЦИЙ
На уровне последнего шейного и
двух первых грудных сегментов спинного мозга расположены группы
преганглионарных симпатических нейронов, получившие название спинно—цилиарного
центра. Они иннервируют гладкие мышцы глазного яблока, мышцы третьего века
у животных одну из мышц верхнего века, глазничную часть круговой мышцы глаза,
мышцу, расширяющую зрачок. Постганглионарные нейроны этого пути лежат в
краниальном шейном симпатическом ганглии. Стимуляция спинно—цилиарного центра,
как и раздражение нервных волокон, сопровождается расширением зрачка (мидриаз),
раскрытием глазной щели, выпячиванием глазного яблока (экзофтальм). Разрушение центра или
перерезка волокон сопровождаются противоположным эффектом — сужением зрачка (миоз),
сужением глазной щели, западением
глазного яблока (эндофтальм).
В первых
пяти грудных сегментах спинного мозга локализованы преганглионарные
симпатические нейроны, имеющие отношение к иннервации сердца и бронхов.
Постганглионарные клетки этого пути лежат в основном в звездчатом ганглии или
реже в узлах пограничного симпатического ствола. Их стимуляция сопровождается
усилением и учащением сердечных сокращений и расширением бронхов.
На всем
протяжении симпатического ядра от первого грудного до начальных поясничных
сегментов расположены скопления клеток, иннервирующих сосуды тела и потовые
железы. Особенностью организации этих скоплений является определенная
топография клеточных тел и в соответствии с этим зон иннервации. Разрушение
клеточных скоплений отдельных сегментов сопровождается исчезновением
сосудистого тонуса, сосудистых реакций и прекращением потоотделения в
определенных областях тела.
В
крестцовой части спинного мозга находятся парасимпатические нейроны, образующие
в совокупности центры дефекации, мочеиспускания, половых рефлексов — эрекции,
эмиссии и эякуляции (см. разд. 14.8.1). При разрушении этих центров названные
рефлексы утрачиваются, возникает нарушение дефекации и мочеиспускания, исчезают
половые рефлексы.
Центры, находящиеся в стволе мозга
(продолговатый мозг, мост, средний мозг), управляют висцеральными функциями
посредством парасимпатических волокон, проходящих в составе блуждающего,
языкоглоточного, лицевого и
глазодвигательного нервов.
В продолговатом мозгу находятся
нервные центры, с помощью которых осуществляются сложные рефлексы, такие
как сосание, жевание, глотание, чихание, кашель, рвота, слезотечение,
слюноотделение. Эти рефлексы имеют в основном защитно—физиологический характер.
Здесь же расположены центры, тормозящие сердечную деятельность, стимулирующие
секрецию желудочных желез, поджелудочной железы, жёлчеобразование и
жёлчевыделение, активирующие моторику желудка и тонкой кишки.
Расположение этих центров
установлено приближенно, а о клеточной организации известно мало.
Ограниченность знаний связана, во—первых, с техническими трудностями, так как
группы нейронов, составляющие центры, небольшие и их трудно различить
нейрофизиологически и морфологически. Во—вторых, клетки, которые управляют
деятельностью какого—либо органа или системы, представляющей собой совокупность
функционально связанных друг с другом органов, не всегда располагаются в одном
месте, рядом. Поэтому понятие «центр» справедливо лишь частично и может
употребляться с некоторыми допущениями.
В ядрах блуждающего нерва
замыкаются рефлексы с аортальной и синокаротидной рефлексогенных зон, а также
рефлексы снижения частоты сердечных сокращений в ответ на раздражение
интероцепторов брюшной полости (рефлекс Гольтца), глазосердечный (рефлекс
Ашнера), дыхательно—сердечный. По волокнам блуждающего нерва следуют импульсы,
регулирующие деятельность дыхательных путей, желудка, тонкой кишки,
поджелудочной железы, жёлчного пузыря, жёлчных протоков. По волокнам лицевого и
языкоглоточного нервов осуществляется регуляция работы слюнных и слезных желез.
Центры зрачкового рефлекса и аккомодации глаза находятся в краниальных
(верхних) холмиках крыши среднего мозга.
Нервные центры продолговатого
мозга оказывают также нисходящие влияния на симпатические и парасимпатические
рефлекторные дуги спинного мозга. Эти влияния могут быть возбуждающими и
тормозными. Особенно отчетливо проявляется влияние нижних отделов ствола мозга
на регуляцию кровообращения. Структуры, осуществляющие эту регуляцию, лежат в
ретикулярной формации продолговатого мозга, их совокупность принято называть сосудодвигательным,
или циркуляторным, центром.
Считают, что именно из
сосудодвигательного центра исходят симпатические тонические влияния на сердце и
сосуды (см. разд. 8.6), в результате чего сердечная деятельность несколько
замедлена, а артерии и артериолы сужены. Это предположение подтверждается тем,
что полная перерезка ствола мозга, проводимая впереди продолговатого мозга,
равно как и выключение практически всех рефлексогенных зон внутренних органов,
мышц, кожи, почти не изменяет уровень артериального давления и его регуляцию. В
отличие от этого у спинальных животных симпатический тонус сосудов снижен и
утрачены приспособительные реакции. Сопряженно с изменением сосудистого тонуса
осуществляются и многие рефлекторные реакции сердца, что объясняют наличием
связей между клеточными скоплениями, регулирующими деятельность сосудов и
деятельность сердца.
Рефлекторные процессы,
происходящие в ядерных образованиях спинного продолговатого, среднего мозга и
моста, находятся под постоянным влиянием гипоталамуса — высшего центра
регуляции висцеральных функций.
5.5.3. Гипоталамические
центры
У позвоночных животных гипоталамус
является главным подкорковым центром интеграции висцеральных процессов. Он
управляет всеми основными гомеостатическими функциями организма. Интегративная
функция гипоталамуса обеспечивается автономными, соматическими и эндокринными
механизмами (они подробно рассмотрены в разд. 2.2, 3.7.3, 6.3). Гипоталамус,
будучи филогенетически наиболее древним образованием, которое существует у всех
хордовых, обнаруживает у них относительно одинаковое строение. Он представляет
собой скопление 32 пар ядер, которые условно разделяются на три группы:
передние, средние и задние.
Чувствительная информация от
внутренних органов и поверхности тела поступает в гипоталамус по восходящим
спинно—бульбарным путям. Одни из них проходят через таламус, другие — через
лимбическую область среднего мозга, третьи следуют по пока еще не полностью
идентифицированным полисинаптическим путям. Кроме того, гипоталамус снабжен и
своими специфическими «входами». В нем имеются высокочувствительные к
изменениям осмотического давления внутренней среды осморецепторы и
чувствительные к изменениям температуры крови терморецепторы.
Эфферентные пути гипоталамуса
полисинаптические. Они связывают его с ретикулярной формацией ствола мозга,
ядрами спинного мозга. Нисходящие влияния гипоталамуса обеспечивают регуляцию
функций главным образом через автономную нервную систему. Вместе с тем важным
компонентом в осуществлении нисходящих влияний гипоталамуса являются и гормоны,
гипофиза. Кроме афферентных и эфферентных связей в гипоталамусе существует комиссуральный
путь. Благодаря ему медиальные гипоталамические ядра одной стороны вступают
в контакт с медиальными и латеральными ядрами другой стороны.
Многочисленные связи гипоталамуса
с другими образованиями мозга способствуют генерализации возбуждений,
возникающих в клетках гипоталамуса. Возбуждение в первую очередь
распространяется на лимбические структуры мозга и через ядра таламуса на
передние отделы коры больших полушарий. Степень распространения восходящих
активирующих влияний гипоталамуса зависит от величины исходного возбуждения
центров гипоталамуса.
Гипоталамус — одна из главных
структур, участвующих в формировании поведенческих реакций организма,
которые необходимы для сохранения постоянства внутренней среды. Стимуляция его
ядер приводит к формированию целенаправленного поведения — пищевого, полового,
агрессивного и т. д. Ему принадлежит и главная роль в возникновении основных
влечений (мотиваций) организма.
Главным источником артериального
кровоснабжения гипоталамических ядер является артериальный круг большого
мозга. Его ветви обеспечивают обильное изолированное кровоснабжение
отдельных групп ядер, капиллярная сеть которых в несколько раз превышает по
густоте кровообеспечение других отделов нервной системы. Капиллярную сеть
гипоталамуса отличает высокая проницаемость для крупномолекулярных соединений.
Рядом с гипоталамусом есть области мозга, в которых гематоэнцефалический барьер
устроен особым образом. В этих местах крупные водорастворимые молекулы гормонов
связываются со специфическими рецепторами и воздействуют на мозг, не проникая в
него (см. раздел 3.27).
Многочисленные нервные и
сосудистые связи между гипоталамусом и гипофизом являются основой
функционального комплекса, называемого гипоталамо—гипофизарной системой.
Главное назначение комплекса состоит в интегрировании нервной и гормональной
регуляции висцеральных функций организма. Со стороны гипоталамуса она
осуществляется двумя путями: парааденогипофизарным (минуя аденогипофиз) и
трансаденогипофизарным (через аденогипофиз).
На высвобождение гормонов передней
доли гипофиза влияют гормоны нейронов гипофизотропной зоны медиальной
области гипоталамуса. Они способны оказывать стимулирующее и тормозное действие
на гипофизарные клетки. В первом случае это так называемые рилизинг—факторы
(либерины), во втором — ингибирующие факторы (статины). Регуляция гипоталамо—гипофизарной
системой висцеральных функций осуществляется по принципу обратной связи. Ее
действие проявляется даже после полного отделения медиальной области
гипоталамуса от других отделов мозга. Роль ЦНС состоит в приспособлении этой
регуляции к внутренним и внешним потребностям организма.
Клетки гипоталамуса избирательно
чувствительны к содержанию тех или иных веществ в крови и при любом изменении
их концентрации приходят в состояние возбуждения. Например, гипоталамические
нейроны чувствительны к малейшим отклонениям рН крови, напряжению O2 и СО3, содержанию
ионов, особенно К+ и Na+. Так, в супраоптическом ядре содержатся клетки,
избирательно чувствительные к изменению осмотического давления крови, в
вентромедиальном ядре — содержанию глюкозы, в переднем гипоталамусе — половых
гормонов.
Следовательно, клетки гипоталамуса
выполняют функции рецепторов, воспринимающих изменение гомеостаза. Они обладают
способностью трансформировать гуморальные изменения внутренней среды в нервный
процесс — биологически окрашенное возбуждение. Однако они могут избирательно
активироваться не только при изменении определенных констант крови, но и
нервными импульсами из соответствующих органов, связанных с данной
потребностью.
Рецепторные клетки работают по триггерному
типу (от англ. trigger—
действие, импульс вызывающее серию явлений). Возбуждение возникает в
них не сразу, как только изменяется какая—либо константа крови, а через
определенный промежуток времени, когда их деполяризация достигнет критического
уровня. Следовательно, нейроны мотивационных центров гипоталамуса отличает
периодичность работы.
В том случае, когда изменение
константы крови поддерживается длительно, деполяризация нейронов поднимается до
критического уровня и состояние возбуждения устанавливается на этом уровне все
время, пока существует изменение константы, вызвавшей развитие процесса
возбуждения. Постоянная импульсная активность этих нейронов исчезает только
тогда, когда устраняется вызвавшее ее раздражение, т. е. нормализуется
содержание того или иного фактора крови. Возбуждение одних клеток гипоталамуса
может возникать периодически через несколько часов, как, например, при
недостатке глюкозы, других — через несколько суток или даже месяцев, как,
например, при изменении содержания половых гормонов.
Разрушение ядер или удаление всего
гипоталамуса сопровождается нарушением гомеостатических функций организма.
Гипоталамус играет ведущую роль в поддержании оптимального уровня метаболизма
(белкового, углеводного, жирового, минерального, водного) и энергии, в
регуляции температурного баланса организма, деятельности сердечно—сосудистой,
пищеварительной, выделительной, дыхательной систем. Под его влиянием находятся
функции эндокринных желез. При возбуждении гипоталамических структур нервный
компонент сложных реакций обязательно дополняется гормональным.
Многочисленные исследования
показали, что стимуляция задних ядер гипоталамуса сопровождается
эффектами, аналогичными раздражению симпатической нервной системы: расширением
зрачков и глазной щели, возрастанием частоты сердечных сокращений, повышением
кровяного давления, торможением моторной активности желудка и кишки,
возрастанием концентрации в , крови адреналина и норадреналина. Задняя область
гипоталамуса оказывает тормозящее влияние на половое развитие. Ее повреждение
приводит также к гипергликемии, а в некоторых случаях к развитию ожирения.
Разрушение задних ядер гипоталамуса сопровождается полной потерей
терморегуляции. Температура тела у этих животных не может поддерживаться, и
животное становится пойкилотермным.
Реакции, возникающие при
возбуждении заднего отдела гипоталамуса и сопровождающиеся активацией
симпатической нервной системы, мобилизацией энергии организма, увеличением
способности к физическим нагрузкам, получили название эрготропных.
Стимуляция группы передних ядер
гипоталамуса характеризуется реакциями, подобными раздражению
парасимпатической нервной системы, сужением зрачков и глазной щели, урежением
частоты сердечных сокращений, снижением величины артериального давления,
усилением моторной активности желудка и кишки, активацией секреции желудочных
желез, возрастанием секреции инсулина и как результат — снижением уровня
глюкозы в крови. Группа передних ядер оказывает стимулирующее влияние на
половое развитие. С ней связан и механизм потери тепла. Разрушение этой области
приводит к нарушению процесса теплоотдачи, в результате чего организм быстро
перегревается. Отношение передних отделов гипоталамуса к соматическим,
висцеральным и эндокринным реакциям в совокупности называют трофотропной
реакцией.
Средняя группа ядер гипоталамуса
обеспечивает главным образом регулирование метаболизма. Изучение
регуляции пищевого поведения показало, что оно осуществляется в результате
реципрокных взаимодействий латерального и вентромедиального гипоталамических
ядер. Активация первого вызывает усиление потребления пищи, а его двустороннее
разрушение сопровождается полным отказом от пищи, вплоть до истощения и гибели
животного. Напротив, повышение активности вентромедиального ядра снижает
уровень пищевой мотивации. При разрушении этого ядра возникает повышение
потребления пищи (гиперфагия), ожирение.
Эти данные позволили расценивать
вентромедиальные ядра как структуры, посредством которых ограничивается прием
пищи, т. е. связанные с насыщением, а латеральные ядра, как структуры, повышающие уровень
пищевой мотивации, т. е. связанные с
голодом.
Вместе с
тем пока еще не удавалось выделить функциональных или структурных скоплении
нейронов, отвечающих за то или иное поведение. Следовательно, клеточные
образования, обеспечивающие формирование целостного поведения из отдельных
реакций, не следует рассматривать как анатомически ограниченные структуры,
известные под названием центр голода и центр насыщения.
Вероятно,
группы клеток гипоталамуса, связанные с выполнением какой—либо функции,
отличаются друг от друга характером афферентных и эфферентных связей,
синаптической организацией и медиаторами. Предполагают, что в нейронных сетях
гипоталамуса заложены многочисленные программы и активация их посредством
сигналов из других отделов мозга или интероцепторов приводит к формированию
необходимых поведенческих и нейрогуморальных реакции.
Изучение
роли гипоталамуса методами раздражения или разрушения его ядер привело к
выводу, что области, ответственные за потребление пищи и воды, по—видимому,
перекрывают друг друга. Полидипсию — повышенное потребление жидкости,
обусловленное патологически усиленной жаждой, — наблюдали при стимуляции
паравентрикулярного ядра гипоталамуса.
С другими
подкорковыми структурами и корой головного мозга гипоталамус находится в
непрерывных циклических взаимодействиях. Благодаря тому что к гипоталамическим
ядрам адресуется нервная и гуморальная сигнализация о различных внутренних
потребностях, они и приобретают значение пускового механизма мотивационных
возбуждений.
Этот
механизм лежит в основе участия гипоталамуса в эмоциональной деятельности
целого организма, что позволило П. К. Анохину и К. В. Судакову высказать
предположение о пейсмекерной роли гипоталамуса в формировании биологических мотиваций. Согласно этому
предположению, гипоталамические пейсмекеры благодаря восходящим активирующим
влияниям определяют энергетическую основу мотивационных возбуждений. Эта
специфика сохраняется в восходящих активирующих влияниях гипоталамуса на всех
уровнях. Тем самым обеспечивается качественное биологическое своеобразие
поведенческих актов. Введение нейротропных веществ специфического действия
может избирательно блокировать различные гипоталамические механизмы,
участвующие в формировании таких состояний организма, как страх, голод, жажда и
т. д.
Гипоталамус
находится под регулирующим влиянием коры головного мозга. Получая информацию об
исходном состоянии организма и окружающей среды, нейроны коры оказывают
нисходящее влияние на все подкорковые структуры, в том числе и гипоталамус,
регулируя уровень их возбуждения. Корковые механизмы подавляют многие эмоции и
первичные возбуждения, формирующиеся с участием гипоталамических ядер. Поэтому
удаление коры нередко приводит к развитию реакций мнимой ярости, выражающейся в
расширении зрачков, пилоэрекции (сокращении мышц, поднимающих волосы),
тахикардии, саливации (слюноотделении), повышении внутричерепного давления и т.
д.
Таким образом, гипоталамус,
обладая хорошо развитой и сложной системой связей, занимает ведущее место в
регуляции многих функций организма и прежде всего в постоянстве внутренней
среды. Под его контролем находится функция автономной нервной системы и
эндокринных желез. Он участвует в регуляции пищевого и полового поведения,
смены сна и бодрствования, эмоциональной деятельности, поддержания температуры
тела и т. д.
Под лимбической системой
принято понимать совокупность функционально связанных между собой образований
древней коры (гиппокамп, грушевидная доля, экторинальная область, параамигдалоидная
кора), старой коры (поясная извилина, пресубикулум) и подкорковых структур
(миндалевидный комплекс, область перегородки, ряд ядер таламуса и гипоталамуса,
а также лимбическая зона среднего мозга). Наряду с управлением висцеральными
функциями лимбическая система участвует в эмоциональном и инстинктивном
(пищевом, половом, оборонительном) поведении. Она оказывает также влияние на
смену фаз сна и бодрствования.
Связь функций лимбической системы с
деятельностью внутренних органов послужила основанием обозначать всю эту
совокупность структур как висцеральный мозг. Однако этот термин лишь
частично отражает функциональное значение системы.
Образования мозга, составляющие
лимбический комплекс (рис. 5.19), относятся к наиболее филогенетически древним
областям и поэтому обнаруживаются у всех позвоночных животных. На начальном
этапе развития позвоночных структуры лимбической системы обеспечивали все
важнейшие реакции организма (ориентировочные, оборонительные, пищевые,
половые), которые формировались на основе первого дистантного чувства — обоняния.
Следовательно, обоняние выступило
в качестве организатора множества целостных функций организма, объединив и их
морфологическую основу — структуру конечного, промежуточного и среднего мозга.
Уже на самых ранних стадиях
эволюции первые акцепторы обонятельной чувствительности — септальная
(миндалевидного тела), амигдалярная
|
|
|
Рис. 5.19
Основные структуры лимбической
системы и связи между ними 1 — клетки
обонятельного эпителия, 2 — обонятельная
луковица, 3 — обонятельный тракт, 4
— ростральная спайка, 5 — мозолистое тело, 6 — поясная извилина, 7 — ростральное ядро таламуса, 8 — терминальная полоска, 9 — свод, 10 — мозговая полоска, 11
— ядра поводка, 12 —
межпоясковое ядро, 13 —
сосцевидное тело, 14 —
миндалевидное тело. |
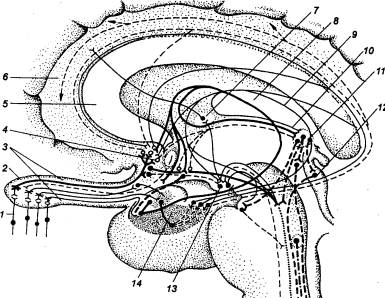
(перегородки) и другие области — были тесно связаны с ядрами
ствола мозга и выполняли наиболее важные интегративные функции, которые обеспечивали
адекватное приспособление организма к условиям окружающей среды. Позже в
результате интенсивного роста новой коры, неостриатума и специфических ядер
таламуса развитие лимбических структур несколько снизилось, но не остановилось.
Они лишь претерпели некоторые топографические изменения и вступили в тесную
связь с более молодыми образованиями мозга, обеспечивая высокоорганизованным
животным более тонкое приспособление к усложняющимся и постепенно меняющимся
условиям обитания.
Лимбическая система обеспечивает
взаимодействие экстероцептивных (обонятельных, слуховых и др.) и
интероцептивных воздействий. Существуют мощные реципрокные связи между
лимбической системой и гипоталамусом, а также височной и лобной долями новой
коры. Височные доли отвечают преимущественно за поступление информации к
миндалине и гиппокампу из зрительной, слуховой и соматосенсорной коры, лобные
доли регулируют деятельность лимбической системы. Характерными для лимбического
комплекса являются и многочисленные цепи возбуждения.
Даже при самой незначительной
стимуляции (электрической, химической и др.) любой из структур лимбической
системы можно обнаружить целый ряд изолированных простых ответов в виде таких
вегетативных проявлений, как саливация, пилоэрекция, изменения работы сердечно—сосудистой,
дыхательной, лимфатической систем, терморегуляции и т. д. Продолжительность
реакций бывает порой значительной, что свидетельствует о включении в работу
эндокринных желез. Часто реакции протекают совместно с координированными
моторными, жевательными, глотательными движениями.
Эти реакции возникают при
стимуляции и разрушении вполне определенных структур. Например, при раздражении
миндалевидного тела, гиппокампа и других областей могут усиливаться или,
напротив, ослабевать оборонительные, пищедобывательные и половые реакции,
нарушаться оценка внутреннего состояния, полезности или вредности внешних
раздражителей, утрачиваться способность к социальному внутригрупповому
поведению.
Наиболее заметно участие
лимбической системы в организации пищевого поведения, реакции утоления жажды.
Так, удаление миндалевидного тела приводит либо к длительному отказу животных
от пищи, либо к гиперфагии. Управляя совокупностью внутренних факторов,
лимбическая система тем самым контролирует эмоциональное состояние животных и
человека, обеспечивая приспособление его видоспецифического поведения,
направленного на приспособление организма. Структуры лимбической системы
связаны с механизмами сна.
Занимая в пределах ЦНС срединное
положение, лимбическая система может быстро включаться практически во все
функции организма, направленные на активное приспособление к условиям
окружающей среды.
В качестве специального отдела
нервной системы мозжечок существует у всех позвоночных животных, ведущих
подвижный образ жизни, хотя только у амфибий впервые встречается его типичное
для высших животных строение.
Уже первые попытки удаления
мозжечка показали его тесную взаимосвязь с двигательной соматической сферой.
Наряду с этим при раздражении мозжечка могут быть воспроизведены практически
все явления, возникающие при возбуждении автономной нервной системы, —
расширение зрачка, сужение сосудов, сокращение мышц, поднимающих волосы,
учащение сердечных сокращений.
|
|
|
Рис. 5.20
Изменение функций различных органов при
раздражении иннервирующих их симпатических и парасимпатических волокон Влияния симпатической нервной
системы обозначены справа, парасимпатической — слева. 1 — сосуды головного мозга, 2 — периферические сосуды, 3 — легкое, 4 — надпочечник, 5 — мочевой пузырь, 6 — внутренние и наружные половые органы, 7 — толстая кишка, 8 — желудок, 9 — сердце, 10 — бронхи, 11 —
слюнная железа, 12 — зрачок. |
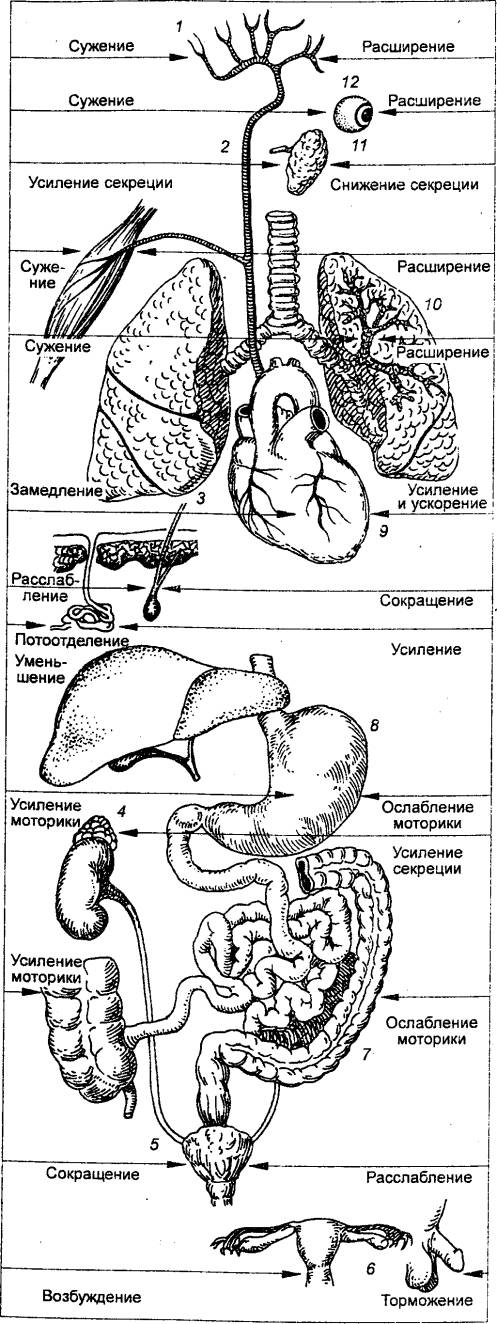
После удаления мозжечка
обнаруживается изменение функционального состояния висцеральных органов. Например,
у лишенных мозжечка животных мышцы, поднимающие волосы, оказываются в состоянии
постоянного сокращения, возникает задержка перистальтики желудка, изменяется
периодическая двигательная деятельность кишки.
Изучение роли мозжечка в регуляции деятельности
сердечно—сосудистой, дыхательной, пищеварительной систем, терморегуляции,
кроветворения, метаболизма и т. д. свидетельствует о том, что мозжечок
благодаря наличию двойного (активирующего и тормозного) механизма действия
способен оказывать стабилизирующее влияние на функции перечисленных
висцеральных систем. Он также играет ведущую роль в корригировании висцеральных
рефлексов.
Взаимодействие мозжечка с высшими
центрами автономной иннервации, эндокринными железами послужило основанием
считать его органом, связанным с регуляторными механизмами автономной нервной
системы.
Во многих отделах ЦНС помимо серого
и белого вещества существует диффузное, скопление клеток, окруженных
многочисленными волокнами. Они идут в разных направлениях и образуют подобие
сети. Это и послужило для немецкого анатома и гистолога О. Дейтерса (1895)
поводом считать эти клетки сетчатым образованием — ретикулярной формацией.
В стволе мозга ретикулярная формация занимает медиальную часть, доходит до
таламуса и гипоталамуса. Ретикулярная формация состоит из нисходящей и
восходящей частей.
Нисходящая часть помимо
скелетной мускулатуры оказывает влияние на функцию внутренних органов. Это
влияние сводится к регуляции деятельности сердца, сосудов, органов дыхания,
пищеварения, выделения. Его проводником на периферию является симпатическая
нервная система (рис. 5.20). Последняя выступает в функциональном единстве с
ретикулярной формацией. В значительной мере ретикулярные влияния сказываются на
функции эндокринных желез и в первую очередь гипофиза. Например, стимуляция
ретикулярных ядер среднего и промежуточного мозга сопровождается усиленной
продукцией гипофизарных, в том числе гонадотропных гормонов. При разрушении
этих ядер у животных возникают эндокринные расстройства, нарушается цикличность
половых процессов.
Восходящая часть
ретикулярной формации передает активирующие влияния в кору больших полушарий.
Ей отводится важная роль в поддержании состояния бодрствования, а также в
механизмах формирования целостных реакций организма.
Таким образом, помимо классической
афферентной системы, называемой специфической, которая несет импульсы в кору
мозга, существует вторая афферентная система — неспецифическая. Она
представлена восходящей частью ретикулярной формации. Окончания первой системы
имеют четкое представительство в определенных проекционных зонах коры и
оказывают локальное действие, тогда как окончания второй расположены во всей
коре и ее действие в основном диффузно.
Влияя на кору, ретикулярная
формация сама испытывает ее постоянное воздействие. Так, в коре существуют
специальные пункты, стимуляция которых вызывает торможение определенных
участков ретикулярной формации. В результате этого в кору не поступают ненужные
сигналы, в том числе и от висцеральных органов. Они блокируются на уровне
ретикулярной формации.
В поддержании активности
ретикулярных механизмов важную роль играют гуморальные раздражители. По
отношению к ним ретикулярная формация обладает высокой чувствительностью. Это
обеспечивает ее участие в регуляции висцеральных функций.
Высшим интегративным центром
регуляции протекающих в организме процессов является кора головного мозга.
Более ста лет назад В. Я. Данилевским было установлено, что
при электрическом раздражении лобных долей возникают изменения сердечной
деятельности и дыхательного ритма. Позже благодаря работам Н. А. Миславского,
В. М. Бехтерева и их последователей накоплено огромное количество фактов,
свидетельствующих о том, что при раздражении или выключении определенных
участков коры больших полушарий можно зарегистрировать изменение
функционального состояния любого органа, иннервируемого автономной нервной
системой (сердце, мышцы, поднимающие волосы, потовые железы, внутренние мышцы
глаза, желудок, кишка, тазовые органы и т. д.). Характерно, что при этом могут
быть зарегистрированы противоположные по знаку изменения висцеральных функций,
как, например, повышение и снижение кровяного давления, усиление и ослабление
кишечной перистальтики и др.
Существует некоторая специализация
отдельных областей коры, при стимуляции которых особенно легко возникают
корковые влияния на определенные процессы. Особенно большое значение имеют лобные
доли коры, из—за чего передние отделы считают высшими центрами
автономной иннервации. В двигательных областях коры находится
представительство тех висцеральных органов и процессов, которые теснейшим
образом связаны с мышечной деятельностью.
Как известно, при мышечной работе
происходит увеличение минутного объема сердца и перераспределение крови в
организме, которая интенсивно снабжает мышцы. Те же сосудистые эффекты легко
воспроизводятся при стимуляции двигательной зоны коры, при этом, кроме того,
изменяется состояние системы терморегуляции, также связанной с мышечной
деятельностью. Благодаря такой организации достигается необходимая для
осуществления нормальной жизнедеятельности интеграция соматических и
висцеральных функций. Эта интеграция характеризуется •высокой точностью.
Возбуждаемые корой висцеральные процессы, так же как и двигательная активность,
находятся в соответствии с индивидуальным опытом животного.
Изменения функционального состояния
автономной нервной системы при стимуляции коры сказываются на всех ее отделах.
Например, расширение зрачка, возникающее при раздражении премоторной зоны коры,
сохраняется у животных после удаления краниального шейного симпатического узла
и полностью исчезает только при последующей церерезке глазодвигательного нерва.
Следовательно, при стимуляции коры возбуждаются симпатические и тормозятся парасимпатические
волокна, иннервирующие радужную оболочку глаза. Бывают и противоположные
эффекты. Влияния эти двусторонние, но наиболее отчетливо они выражены на
контралатеральной стороне тела.
Многочисленными опытами доказано,
что информация о работе внутренних органов при определенных условиях может
достигать коры больших полушарий мозга. Данные, полученные В. Н. Черниговским
при изучении интероцепции, подтвердили выдвинутое И. П. Павловым положение о
существовании особого интероцептивного анализатора, воспринимающего
раздражения из внутренней среды организма.
Используя метод условных рефлексов,
К. М. Быков и его ученики доказали, что раздражение рецепторов практически всех
внутренних органов может оказывать мощное влияние на процессы высшей нервной
деятельности. Установлено также, что на основе раздражения рецепторов
внутренних органов могут быть выработаны интероцептивные условные рефлексы.
Возможности образования висцеральных условных рефлексов наряду с вегетативными
сдвигами при различных эмоциях были положены К. М. Быковым в основу концепции
существования кортико—висцералъных отношений.
Согласно этой концепции, кора
больших полушарий моделирует деятельность подкорковых структур, имеющих
непосредственное отношение к регуляции внутренней среды организма. Существенную
роль при этом играют прямые и опосредованные через ретикулярную формацию связи
коры с гипоталамусом и гипофизом. Часть волокон, по которым осуществляется
корковый контроль висцеральных функций, проходит в составе пирамидных путей.
Основной функцией автономной
нервной системы является регулирование процессов жизнедеятельности
органов тела, согласование и приспособление их работы к общим нуждам и
потребностям организма.
Автономная нервная система представляет
собой совокупность нервных волокон периферических узлов, состоящих из нервных
клеток, центров спинного мозга и ствола мозга. Автономная и
соматическая рефлекторные дуги построены по одному и тому же плану и содержат
чувствительное, ассоциативное и эффективное звенья. Различие их заключается в
том, что эфферентные клетки автономной нервной системы лежат в ганглиях вне
ЦНС.
Таким образом, автономная нервная
система по составу нейронных звеньев дуги, нисколько не отличающемуся от
соматической, совершенно не характеризуется чисто эфферентной природой, как это
ей приписывали до недавнего времени.
Выдвижение в качестве отличительной черты автономной нервной системы
двухнейронности ее центробежных периферических проводников также лишено
основания. Доказательством тому является следующее. Преганглионарные
симпатические нейроны боковых рогов спинного мозга соответствуют не моторным, а
ассоциативным (вставочным) элементам соматической дуги. Моторные же нейроны
соматической нервной системы соответствуют эффекторным нейронам периферических
автономных ганглиев. Исходя из этого синапс между пре— и постганглионарными
волокнами в автономном ганглии аналогичен внутримозговому синапсу между
ассоциативным и двигательным соматическим нейронами и отличается от него только
топографически: он вынесен на периферию.
Автономная нервная система состоит
из трех частей: симпатической, парасимпатической и метасимпатической.
Анатомически симпатическая и парасимпатическая части различаются местами выхода
из ЦНС. Первая выходит из грудо—поясничного отдела спинного мозга, вторая — из
среднего мозга, продолговатого мозга и крестцовой части спинного мозга.
Тело преганглионарного нейрона
располагается в сером веществе ЦНС, постганглионарного симпатического — в пара—
и превертебральных ганглиях, парасимпатического — в непосредственной близости
от эффекторного органа. Весь рефлекторный аппарат метасимпатической нервной
системы находится в ганглиях и нервных сплетениях стенок внутренних органов.
Большинство висцеральных органов имеет тройную — симпатическую,
парасимпатическую и метасимпатическую — иннервацию.
Симпатическая часть
возбуждается относительно диффузно, парасимпатическая — характеризуется
более ограниченными по областям распространения рефлекторными реакциями. Метасимпатическая
часть является высокоорганизованным функциональным ансамблем, наделенным
большой степенью автономии и способностью к интегративным процессам.
Симпатический отдел — это система
тревоги, система «защиты», система мобилизации резервов, необходимая для
взаимодействия организма со средой. Эта мобилизация достигается
генерализованным вовлечением в реакцию многих органов и систем. Вероятно,
поэтому симпатические ганглии расположены на большом расстоянии от
иннервируемых органов и обладают способностью к мультипликации импульсов, что и
обеспечивает быструю генерализацию воздействия. Более медленный, но также
генерализованный процесс возникает при выбросе в кровь адреналина, образно
представляемого в качестве «жидкой симпатической системы». Симпатические
импульсы активируют деятельность мозга, мобилизуют защитные реакции: процессы
терморегуляции, механизмы свертывания крови, иммунные реакции. Возбуждение
симпатической нервной системы — обязательное условие эмоционального состояния и
напряжения, оно является также начальным звеном гормональных реакций стресса.
В отличие от этого
парасимпатический и метасимпатический отделы представляют системы текущей
регуляции физиологических процессов в организме, обеспечивающих
гомеостатическое состояние.
Функциональные свойства нейронов метасимпатической
нервной системы сопоставимы со свойствами клеток ядерных образований мозга.
Первоначальное предположение о функции метасимпатических нейронов как простом
переключении информации, получаемой по волокнам из парасимпатического отдела
автономной нервной системы, не соответствует действительности.
Метасимпатические нейроны, конечно, получают сигналы, передаваемые из ЦНС по
парасимпатическим и симпатическим волокнам, однако это лишь один из способов
ввода информации. Эта система имеет собственную интегративную цепь для
обработки информации, поступающей не только по симпатическим и
парасимпатическим путям, но и от рецепторов стенок полых внутренних органов. По
существующей концепции метасимпатическую нервную систему рассматривают как
независимую систему, которая программирует и координирует двигательную
активность и другие функции висцеральных органов, такие как секреция,
всасывание. Под ее влиянием находятся местные эндокринные элементы и локальный
кровоток, иммунные процессы.
Тонкая координация деятельности
всех трех частей автономной нервной системы осуществляется надсегментарными
центрами при участии коры больших полушарий головного мозга. В гипоталамической
области, этом сложно организованном отделе промежуточного мозга, находятся
ядра, имеющие непосредственное отношение к деятельности автономной нервной
системы.
Тела чувствительных клеток дуги
автономного рефлекса могут располагаться в спинальных и периферических
ганглиях. В последнем случае один из отростков клетки направляется в спинной
мозг, другой — во внутренние органы. Помимо участия в спинальной дуге рефлекса
такие нейроны имеют прямое отношение к местным (периферическим) рефлекторным
процессам. Чувствительные волокна в боковых рогах спинного мозга образуют
моносинаптические контакты с преганглионарными нейронами, их связь носит
полисинаптический характер.
При малой интенсивности раздражения
рецептора в реакцию, как правило, вовлекаются лишь местные рефлекторные дуги
ганглионарного уровня, что сопровождается соответствующей реакцией
исполнительного органа (сосудистые эффекты, изменение перистальтики и др.). С
повышением интенсивности афферентного потока последовательно активируются
сегментарная, проприоспинальная система, а также восходящие пути, которые
вовлекают супраспинальные структуры, связанные с висцеральными функциями. В
итоге возникают обширные, или генерализованные, ответные реакции,
сопровождающиеся иногда субъективными ощущениями.
Синаптические связи в автономных
ганглиях сложны и разнообразны. На одном эффекторном нейроне может оканчиваться
несколько отличающихся по функциональным характеристикам преганглионарных
волокон. Синаптическая задержка в разных ганглиях составляет от 1 до 10 мс. Она
особенно продолжительна в периферических рефлекторных путях.
Возбуждение преганглионарных
нервных волокон приводит к выделению из них ацетилхолина. Скорость
синтеза передатчика обеспечивает не ограниченную во времени передачу активности
при естественной частоте ее следования. В мембранах пресинаптических и
постсинаптических структур независимо от того, к какой системе они принадлежат
— симпатической, парасимпатической или метасимпатической, содержатся М—
и Н—холинорецепторы. Одной и той же клетке могут принадлежать оба вида
рецепторов.
В естественных условиях по
преганглионарным волокнам непрерывно поступает центробежная тоническая
импульсация. Она может следовать либо в виде отдельных импульсов, либо в виде
синхронных залпов разрядов, вызывающих в постганглионарном нейроне множество
локальных возбуждений.
Постганглионарная клетка дуги
автономного рефлекса во многих отношениях напоминает мотонейрон. Это низкий
предельный ритм импульсации длительная следовая гиперполяризация, ответы на
преганглионарный залп всего одним импульсом и т. п. Сходство еще более
усиливается совпадениями функционального назначения, так как обе клетки
передают сигналы исполнительным органам, минуя промежуточные инстанции.
В автономной нервной системе
известны три вида синоптической передачи: электрическая, химическая,
смешанная. Электрические синапсы располагаются преимущественно там, где
необходима быстрая передача возбуждения, например в ресничном ганглии птиц.
Основную же массу составляют синапсы с химической передачей. Сейчас в
автономной нервной системе насчитывается более десяти типов нервных клеток,
которые продуцируют разные медиаторы — ацетилхолин, норадреналин, серотонин и
другие биогенные амины, АТФ, различные нейропептиды. В связи с этим их принято
называть холинергическим, адренергическим, серотонинергическим, пуринергическим
нейронами.
Рассматривая вопрос о локализации
медиаторных систем в эффекторных путях автономной нервной системы, можно прийти
к следующей простой схеме. В симпатической нервной системе передача с
преганглионарных на постганглионарные нейроны осуществляется в основном с помощью
ацетилхолина, на эффектор — посредством норадреналина. В
парасимпатической нервной системе передатчиком возбуждения в обоих звеньях
эфферентного пути является ацетилхолин. В метасимпатической нервной системе
взаимодействуют оба эти медиатора. Помимо того, медиация в эффекторном нейроне
осуществляется преимущественно с помощью серотонина и АТФ.
Существуют и иные варианты. В
ганглионарной межнейронной передаче и автономной нервно—мышечной передаче могут
участвовать и другие медиаторы, а также биологически активные вещества типа
пептидов. Сейчас с деятельностью автономной нервной системы связывают более 20
классических и предполагаемых медиаторов.
Багаев В. А., Ноздрачев А.
Д., Пантелеев С. С. Ваговагальная рефлекторная дуга. Элементы структурно—функциональной
организации. СПб., 1997.
Баклаваджян О. Г. Центральные механизмы
гомеостаза // Частная физиология нервной системы /Отв. ред. П. Г. Костюк. Л„
1983. С. 216—312.
Баклаваджян
О. Г.
Нейронная организация гипоталамо—висцеральной рефлекторной дуги. Л., 1988.
Блум Ф.,
Лейзерсон Ф., Хофстедтер Л. Мозг, разум и поведение. М., 1988.
Вейн А. М. Вегетативная нервная система
(руководство для врача). М., 1998.
Говырин В.
А.
Трофическая функция симпатических нервов сердца и скелетных мышц. Л., 1967.
Глазачев О.
С.
Вегетативная нервная система. Принцип строения. Функция. Методы
исследования.М., 1995.
Костюк П.
Г., Преображенский Н. Н. Механизмы интеграции висцеральных и соматических афферентных сигналов.
Л.,1975. Ноздрачев А. Д. Вегетативная рефлекторная дуга. Л., 1978.
Ноздрачев А.
Д.
Физиология вегетативной нервной системы. Л., 1983.
Ноздрачев А.
Д., Пушкарев Ю. П. Характеристика медиаторных превращений. Л., 1980.
Ноздрачев А.
Д., Чернышева М. П. Висцеральные рефлексы. Л., 1989.
Ноздрачев А.
Д.. Чумасов Е. И. Периферическая нервная система. СПб., 1999.
Ноздрачев А.
Д., Янцев А. В. Автономная передача. СПб., 1995. Нормальная физиология: Курс физиологии
функциональных систем / Под ред. К. В. Судакова.М., 1999.
Скок В. И. Физиология вегетативных
ганглиев. Л., 1970.
Скок В. И.,
Иванов А. Я.
Естественная активность вегетативных ганглиев. Киев, 1989.
Физиология
человека. В 2 т. / Под ред. В. М. Покровского, Г. Ф.Коротько. М., 1997.
Физиология
человека. В 3 т. / Под ред. Р. Шмидта, Г. Тевса. М., 1996.
Харди Р. Гомеостаз. М., 1986.
Черниговский
В. Н.
Нейрофизиологический анализ кортико—висцеральной рефлекторной дуги.Л., 1967.
Шеперд Г. Нейробиология. В 2 т. М.,
1987.
Шмидт—Ниельсен К. Физиология животных.
Приспособление и среда. В 2 кн. М., 1982. Кн. 2.
Эккерт Р., Рэнделл Д., Огастин Дж. Физиология
животных. Механизм и адаптация: В 2 т. М., 1991.