3 ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
VI в. до н. э. — Пифагор
Самосский (Др. Греция) считал мозг средоточием психической деятельности. V—IV вв. до н.
э. — Гиппократ (Др.
Греция) описал четыре типа темперамента. V—I вв. до н.
э. — Демокрит (Др.
Греция) и Лукреций (Др. Рим) считали мозг седалищем души. IV в. до н. э. — Аристотель (Др.
Греция) создал известный труд по психологии — трактат «О душе».
1012—1024 — Авиценна (Абу Али ибн Сина,
Центральная Азия, Иран) в «Каноне врачебной науки» дал описание нервов
черепномозговых и спинномозговых, чувствительных и двигательных. Считал мозг
местом пребывания душевной силы и источником ее действия.
1637 — Р. Декарт (R. Descartes, Франция) дал первое описание
двигательного рефлекса.
1784 — И. Прохаска (J. Prochaska, Чехия) развил представление о
рефлексе и предложил этот термин. Описал рефлекторную дугу.
1811 и 1822 — Ч. Белл (Ch. Bell, Великобритания) и Ф. Мажанди (F. Magendie, Франция) открыли порядок
распределения чувствительных и двигательных нервных волокон между дорсальными и
вентральными корешками спинного мозга (закон Белла—Мажанди).
1832 — М. Голл и
И. Мюллер (М. Goll, J. Muller, Германия) изложили учение о рефлексе.
1838 — Т. Шванн (Th. Schwann, Германия) описал строение оболочки
нервного волокна (шванновская оболочка).
1856 — Р. Вирхов
(R. Virchow, Германия) — открыл нейроглию.
1863 — И. М. Сеченов (Россия) в работе
«Рефлексы головного мозга» описал явление центрального торможения и
распространил принцип рефлекторной реакции на психическую деятельность и
поведение человека.
1865 — О. Дейтерс (О. Deiters, Германия) описал отростки нейронов
— аксон и дендриты.
1874 — В. А. Бец (Россия) описал
гигантские пирамидные клетки в коре больших полушарий головного мозга,
непосредственно управляющие мотонейронами скелетных мышц.
1874 — В. Вундт (W. Wundt, Германия) основал первую лабораторию
экспериментальной психологии, ввел психометрию — количественный метод изучения
психики.
1885 — П. Эрлих (Р. Ehrlich, Германия) впервые высказал мысль о существовании барьера
между кровью и мозгом.
1891 — Г. Вальдейер (Н. Waldeyer, Германия) заявил о применимости
клеточной теории к нервной системе и предложил термин «нейрон».
1895 — 3. Фрейд (Z. Freud, Австро—Венгрия) заложил основы
психоанализа.
1897 — Ч.
Шеррингтон (Ch. Sherrington, Великобритания) ввел понятие «синапс».
1902 — в докладе из лаборатории И. П.
Павлова впервые заявлено об открытии условных рефлексов.
1905 — А. Бине и
Т. Симон (A. Binet, Th. Simon, Франция) создали первый тест для оценки интеллекта (впоследствии IQ — «коэффициент
интеллектуальности»).
1906 — К. Гольджи
(С. Golgi, Италия) и С. Рамон—и—Кахаль (S. Ramon у Cajal, Испания) — Нобелевская премия за
доказательство клеточного строения нервной системы.
1909 — К. Бродман (К. Brodmann, Германия) описал в коре больших
полушарий головного мозга 11 областей, включающих в себя 52
цитоархитектонических поля.
1921 — К. Юнг (К. Jung, Швейцария) в основу типологии
характеров положил доминирующую психическую функцию индивида. Ввел понятия об
интра— и экстраверсии.
1923 — А. А.
Ухтомский (СССР) создал учение о доминанте.
1924 — Р. Магнус
и А. Де Клейн (R. Magnus, A. De Kleijn, Нидерланды) ввели понятие о
статических и статокинетических рефлексах ствола мозга.
1925 — В.
Правдич—Неминский (СССР) зарегистрировал электрические процессы в мозгу
животного. 1929 — Л. С. Штерн (СССР)
ввела понятие о гистогематических барьерах.
1929 — Г. Бергер
(Н. Berger, Германия) зарегистрировал электрические процессы в мозгу человека.
1920—1930—е — Л. С. Выготский (СССР) исследовал
развитие мышления и речи.
1932 — Ч. Шеррингтон и Э. Эдриан (Е. Adrian, Великобритания) — Нобелевская
премия за открытие роли рецепторов нервов, передачи информации в виде
электрических импульсов.
1936 — О. Лёви
(О. Loewi, Австрия) и Г. Дейл (Н. Dale, Великобритания) — Нобелевская премия за открытие
механизма синаптической передачи.
1937 — Дж. Папес
(J. Papez, США) описал функции «круга
Папеса», включающего в себя гиппокамп, маммилярное тело, часть таламуса, кору
поясной извилины и другие структуры мозга.
1938 — К. Хейманс (С. Heymans, Бельгия) — Нобелевская премия за открытие роли синусного и
аортального механизмов в регуляции дыхания.
1930—е — У. Пенфилд (W. Penfield, Канада) обнаружил правильную
пространственную проекцию скелетных мышц на двигательную область коры больших
полушарий головного мозга.
1938 — Б. Скиннер (В. Skinner, США) предложил камеру для
выработки инструментальных рефлексов у крыс (камера Скиннера).
1941 — Н. А. Бернштейн (СССР) в трудах
по физиологии движений создал схему рефлекторного кольца.
1943 — Д. Ллойд (D. Lloyd, Великобритания) установил, что
рефлекс на растяжение скелетной мышцы является моносинаптическим.
1944 — Г. Мэгун
(Н. Magoun, США) установил, что электрическое раздражение ретикулярной формации
ствола мозга может либо активировать, либо тормозить двигательные рефлексы.
1944 — Дж. Эрлангер и Г. Гассер (J. Erianger, Н. Gasser, США) — Нобелевская премия за
открытие функциональных различий между нервными волокнами различных типов.
1949— В. Хесс (W. Hess, Швейцария)— Нобелевская премия за
открытие гипоталамических центров регуляции вегетативных функций.
1950 — У. Пенфилд обнаружил
соматотопическую локализацию моторных функций в коре больших полушарий
головного мозга.
1952 — П. Маклин (Р. MacLean, США) развил идеи Папеса и ввел
понятие «лимбическая система».
1957 — В. Маунткасл (V. Mountcastle, США) показал, что соматосенсорная
кора больших полушарий головного мозга организована в элементарные
функциональные единицы — колонки.
1957 — Ч. Осгуд (Ch. Osgood, США) разработал метод
семантического дифференциала.
1963 — Дж. Эклс (J. Eccles, Австралия), А. Ходжкин (A. Hodgkin, Великобритания) и А. Хаксли (A. Huxley, Великобритания) — Нобелевская
премия за открытие ионных механизмов передачи возбуждения по нервному волокну.
1965 — Э. Хеннеман (Е. Henneman, ФРГ) описал законы вовлечения в
возбуждение мотонейронов двигательных ядер в зависимости от их размеров и
возбудимости.
1960—е — А. Р. Лурия (СССР) применил психологические методы для анализа
поражений мозга.
1970 — Б. Кац (В. Katz, Великобритания), У. фон Эйлер (U. v. Euler, Швеция) и Дж. Аксельрод (J. Axelrod, США) — Нобелевская премия за
открытие роли норадреналина в синаптической передаче.
1970 — Р. Гранит (R. Granit, Швеция) опубликовал классическую
монографию о физиологических механизмах регуляции движений.
1973 — К. фон Фриш (К. v. Frisch, ФРГ), К. Лоренц (К. Lorenz, Австрия) и Н. Тинберген (N. Tinbergen, Великобритания) — Нобелевская
премия за создание этологии (науки о поведении).
1975 — Дж. Хьюз и Г. Костерлиц (J. Huges, H. Kosterlitz, Великобритания) обнаружили в мозгу
энкефалины — медиаторы опиоидной системы.
1977 — Р. Гиймен и Э. Шалли (R. Guillemin, A. Schally, США) — Нобелевская премия за
открытие тиролиберина — первого из рилизинг—факторов гипоталамуса.
1977 — Р. Ялоу ( R. Yalow, США) — Нобелевская премия за развитие
методов радиоиммунологического исследования (RIA) пептидных гормонов.
1979 — А. Кормак (A. Cormack, США) и Г. Хаунсфилд (G. Hounsfield, Великобритания) — Нобелевская
премия за разработку метода компьютерной томографии.
1981 — Р. Сперри (R. Sperry, США) — Нобелевская премия за
открытие функциональной специализации полушарий мозга.
2000 — А. Карлссон (A. Carlsson, Швеция), П. Грингард и Э. Кэндел
(Р. Greengard, Е. Kandel, СИТА) — Нобелевская премия за открытия в области
передачи сигналов в нервной системе.
3.1. ЭТАПЫ РАЗВИТИЯ НЕРВНОЙ СИСТЕМЫ
Появление многоклеточных организмов
явилось первичным стимулом для дифференциации систем связи, которые
обеспечивают целостность реакций организма, взаимодействие между его тканями и
органами. Это взаимодействие может осуществляться гуморальным путем посредством
поступления гормонов и продуктов метаболизма в кровь, лимфу и тканевую жидкость
и за счет функции нервной системы, которая обеспечивает быструю передачу
возбуждения, адресованного к вполне определенным мишеням.
3.1.1. Нервная система
беспозвоночных животных
Нервная система как специализированная система интеграции на
пути структурного и функционального развития проходит через несколько этапов,
которые у первично— и вторичноротых животных могут характеризоваться чертами
параллелизма и филогенетической пластичностью выбора.
Среди беспозвоночных наиболее примитивный тип нервной
системы в виде диффузной нервной сети встречается у кишечнополостных
(см. рис. 1.2). Их нервная сеть представляет собой скопление мультиполярных и
биполярных нейронов, отростки которых могут перекрещиваться, прилегать друг к
другу и лишены функциональной дифференциации на аксоны и дендриты. Диффузная
нервная сеть не разделена на центральный и периферический отделы и может быть
локализована в эктодерме и энтодерме.
Эпидермальные нервные сплетения, напоминающие нервные сети
кишечнополостных, могут быть обнаружены и у более высоко организованных
беспозвоночных (плоские и кольчатые черви), однако здесь они занимают
подчиненное положение по отношению к ЦНС, которая выделяется как
самостоятельный отдел.
В качестве примера такой централизации и концентрации
нервных элементов можно привести ортогональную (от греч. orthogonios — прямоугольный) нервную систему
плоских червей. Ортогон высших турбеллярий представляет собой упорядоченную
структуру, которая состоит из ассоциативных и двигательных клеток, формирующих
вместе несколько пар продольных тяжей, или стволов, соединенных большим числом
поперечных и кольцевых комиссуральных стволов. Концентрация нервных элементов
сопровождается их погружением в глубь тела.
Плоские черви являются билатерально симметричными животными
с четко выраженной продольной осью тела. Движение свободноживущих форм
осуществляется преимущественно в сторону головного конца, где концентрируются
рецепторы, сигнализирующие о приближении источника раздражения. К числу таких
рецепторов турбеллярий относятся пигментные глазки, обонятельные ямки,
статоцист, чувствительные клетки покровов (см. разд. 4.2.1 и 4.5), наличие
которых способствует концентрации нервной ткани на переднем конце тела. Этот
процесс приводит к формированию головного ганглия, который, по меткому
выражению Ч. Шеррингтона, можно рассматривать как ганглиозную надстройку над
системами рецепции на расстоянии.
Ганглионизация нервных элементов получает дальнейшее развитие у
высших беспозвоночных, кольчатых червей, моллюсков и членистоногих. У
большинства кольчатых червей брюшные стволы ганглионизированы таким образом,
что в каждом сегменте тела формируется по одной паре ганглиев, соединенных
коннективами с другой парой, расположенной в соседнем сегменте.
Ганглии одного сегмента у примитивных аннелид соединены
между собой поперечными комиссурами, и это приводит к образованию лестничной
нервной системы. В более продвинутых отрядах кольчатых червей
наблюдается тенденция к сближению брюшных стволов вплоть до полного слияния
ганглиев правой и левой сторон и перехода от лестничной цепочечной нервной
системе. Идентичный, цепочечный тип строения нервной системы существует у
членистоногих с различной выраженностью концентрации нервных элементов, которая
может осуществляться не только за счет слияния соседних ганглиев одного
сегмента, но и при слиянии последовательных ганглиев различных сегментов.
Эволюция нервной системы беспозвоночных идет не только по
пути концентрации нервных элементов, но и в направлении усложнения структурных
взаимоотношений в пределах ганглиев. Не случайно брюшную нервную цепочку
сравнивают со спинным мозгом позвоночных животных. Как и в спинном
мозгу, в ганглиях обнаруживается поверхностное расположение проводящих путей,
дифференциация нейропиля на моторную, чувствительную и ассоциативные области.
Это сходство, являющееся примером параллелелизма в эволюции тканевых структур,
не исключает, однако, своеобразия анатомической организации. Так, например,
расположение туловищного мозга кольчатых червей и членистоногих на брюшной
стороне тела обусловило локализацию моторного нейропиля на дорсальной стороне
ганглия, а не на вентральной, как это имеет место у позвоночных животных.
Процесс ганглионизации у беспозвоночных может привести к
формированию нервной системы разбросанно—узлового типа, которая
встречается у моллюсков. В пределах этого многочисленного типа имеются
филогенетически примитивные формы с нервной системой, сопоставимой с ортогоном
плоских червей (боконервные моллюски), и продвинутые классы (головоногие
моллюски), у которых слившиеся ганглии формируют дифференцированный на отделы
мозг, защищенный гематоэнцефалическим барьером (см. разд. 3.27).
Прогрессивное развитие мозга у головоногих моллюсков и
насекомых создает предпосылку для возникновения своеобразной иерархии командных
систем управления поведением. Низший уровень интеграции в сегментарных
ганглиях насекомых и в подглоточной массе мозга моллюсков служит основой для
автономной деятельности и координации элементарных двигательных актов. В то же
время мозг представляет собой следующий, более высокий уровень интеграции,
где могут осуществляться межанализаторный синтез и оценка биологической
значимости информации. На основе этих процессов формируются нисходящие команды,
обеспечивающие вариантность запуска нейронов сегментарных центров. Очевидно,
взаимодействие двух уровней интеграции лежит в основе пластичности поведения
высших беспозвоночных, включающего врожденные и приобретенные реакции.
В целом, говоря об эволюции нервной системы беспозвоночных,
было бы упрощением представлять ее как линейный процесс. Факты, полученные в
нейроонтогенетических исследованиях беспозвоночных, позволяют допустить
множественное (полигёнетическое) происхождение нервной ткани беспозвоночных.
Следовательно, эволюция нервной системы беспозвоночных могла идти широким
фронтом от нескольких источников с изначальным многообразием.
На ранних этапах филогенетического развития сформировался второй
ствол эволюционного древа, который дал начало иглокожим и хордовым.
Основным критерием для выделения типа хордовых является наличие хорды,
глоточных жаберных щелей и дорсального нервного тяжа — нервной трубки,
представляющей собой производное наружного зародышевого листка эктодермы. Трубчатый
тип нервной системы позвоночных по основным принципам организации отличен
от ганглионарного, или узлового, типа нервной системы высших беспозвоночных .
3.1.2. Нервная система позвоночных животных
Нервная система позвоночных закладывается в виде сплошной
нервной трубки, которая в процессе онто— и филогенеза дифференцируется на
различные отделы и является также источником периферических симпатических,
парасимпатических и метасимпатических нервных узлов. У наиболее древних
хордовых (бесчерепных) головной мозг отсутствует, и нервная трубка представлена
в малодифференцированном состоянии.
Согласно представлениям Л. А. Орбели, С. Херрика, А. И.
Карамяна, этот критический этап развития центральной нервной системы
обозначается как спинальный. Нервная трубка современного бесчерепного
(ланцетника), как и спинной мозг более высокоорганизованных позвоночных, имеет
метамерное строение и состоит из 62—64 сегментов, в центре которых проходит спинномозговой
канал. От каждого сегмента отходят брюшные (двигательные) и спинные
(чувствительные) корешки, которые не образуют смешанных нервов, а идут в виде
отдельных стволов. В головных и хвостовых отделах нервной трубки локализованы
гигантские клетки Родэ, толстые аксоны которых образуют проводниковый аппарат.
С клетками Родэ связаны светочувствительные глазки Гесса, возбуждение которых
вызывает отрицательный фототаксис (см. разд. 4.8.3).
В головной части нервной трубки ланцетника находятся крупные
ганглиозные клетки Овсянникова, имеющие синаптические контакты с биполярными
чувствительными клетками обонятельной ямки. В последнее время в головной части
нервной трубки идентифицированы нейросекреторные клетки, напоминающие
гипофизарную систему высших позвоночных. Однако анализ восприятия и простых
форм обучения ланцетника показывает, что на данном этапе развития ЦНС
функционирует по принципу эквипотенциальности, и утверждение о специфике
головного отдела нервной трубки не имеет достаточных оснований.
В ходе дальнейшей эволюции наблюдается перемещение некоторых
функций и систем интеграции из спинного мозга в головной — процесс
энцефализации, который был рассмотрен выше на примере беспозвоночных
животных. В период филогенетического развития от уровня бесчерепных до уровня
круглоротых формируется головной мозг как надстройка над системами
дистантной рецепции.
Исследование ЦНС современных круглоротых показывает, что их
головной мозг в зачаточном состоянии содержит все основные структурные
элементы. Развитие вестибулолатеральной системы, связанной с полукружными
каналами и рецепторами боковой линии, возникновение ядер блуждающего нерва и
дыхательного центра создают основу для формирования заднего мозга.
Развитие дистантной зрительной рецепции дает толчок к
закладке среднего мозга. На дорсальной поверхности нервной трубки
развивается зрительный рефлекторный центр — крыша среднего мозга, куда приходят
волокна зрительного нерва. И наконец, развитие обонятельных рецепторов
способствует формированию переднего, или конечного, мозга, к которому
примыкает слаборазвитый промежуточный мозг.
Указанная выше направленность процесса энцефализации
согласуется с ходом онтогенетического развития мозга у круглоротых. В процессе
эмбриогенеза головные отделы нервной трубки дают начало трем мозговым пузырям.
Из переднего пузыря формируется конечный и промежуточный мозг, средний
пузырь дифференцируется в средний мозг, а из заднего пузыря образуются продолговатый
мозг и мозжечок. Сходный план онтогенетического развития мозга
сохраняется и у других классов позвоночных.
Нейрофизиологические исследования мозга круглоротых
показывают, что его главный интегративный уровень сосредоточен в среднем и
продолговатом мозгу, т. е. на данном этапе развития ЦНС доминирует бульбомезенцефальная
система интеграции, пришедшая на смену спинальной.
Передний мозг круглоротых длительное
время считали чисто обонятельным. Однако исследования недавнего времени
показали, что обонятельные входы в передний мозг не являются единственными, а
дополняются сенсорными входами других модальностей. Очевидно, уже на ранних
этапах филогенеза позвоночных передний мозг начинает участвовать в переработке
информации и управлении поведением. Вместе с тем энцефализация как
магистральное направление развития мозга не исключает эволюционных преобразований
в спинном мозгу круглоротых. В отличие от бесчерепных нейроны кожной
чувствительности выделяются из спинного мозга и концентрируются в
спинномозговой ганглий. Наблюдается совершенствование проводниковой части
спинного мозга. Проводящие волокна боковых столбов имеют контакты с мощной
дендритной сетью мотонейронов. Формируются нисходящие связи головного мозга со
спинным через мюллеровские волокна — гигантские аксоны клеток, лежащих в
среднем и продолговатом мозгу.
Появление более сложных форм
двигательного поведения у позвоночных сопряжено с совершенствованием
организации спинного мозга. Так, например, переход от стереотипных ундулирующих
движений круглоротых к локомоции с помощью плавников у хрящевых рыб (акулы,
скаты) связан с разделением кожной и мышечно—суставной (проприоцептивной)
чувствительности (см. разд. 4.3). В спинальных ганглиях появляются
специализированные нейроны для выполнения этих функций.
В эфферентной части спинного мозга
хрящевых рыб также наблюдаются прогрессивные преобразования. Укорачивается путь
моторных аксонов внутри спинного мозга, происходит дальнейшая дифференциация
его проводящих путей. Восходящие пути боковых столбов у хрящевых рыб доходят до
продолговатого мозга и мозжечка. Вместе с тем восходящие пути задних столбов
спинного мозга еще не дифференцированы и состоят из коротких звеньев.
Нисходящие пути спинного мозга у
хрящевых рыб представлены развитым ретикулярно—спинномозговым путем и путями,
соединяющими вестибулолатеральную систему и мозжечок со спинным мозгом
(преддверно—спинномозговой и мозжечково—спинномозговой пути).
Одновременно в продолговатом мозгу
наблюдается усложнение системы ядер вестибулолатеральной зоны. Этот процесс
сопряжен с дальнейшей дифференциацией органов боковой линии и с появлением в
лабиринте третьего (наружного) полукружного канала в дополнение к переднему и
заднему.
Развитие общей двигательной
координации у хрящевых связано с интенсивным развитием мозжечка (рис.
3.1). Массивный мозжечок акулы имеет двусторонние связи со спинным,
продолговатым мозгом и покрышкой среднего мозга. Функционально он разделяется
на части: старый мозжечок (археоцеребеллум), связанный с вестибулолатеральной
системой, и древний мозжечок (палеоцеребеллум), включенный в систему анализа
проприоцептивной чувствительности. Существенным моментом структурной
организации мозжечка хрящевых рыб является его многослойность. В сером веществе
мозжечка акулы идентифицированы молекулярный слой, слой клеток Пуркинье и
зернистый слой.
Другой многослойной структурой
стволовой части мозга хрящевых рыб является крыша среднего мозга, куда
подходят афференты различных модальностей (зрительные, соматические). Сама
морфологическая организация среднего мозга свидетельствует о его важной роли в
интегративных процессах на данном уровне филогенетического развития.
В промежуточном мозгу хрящевых рыб происходит дифференциация
гипоталамуса, который является наиболее древним образованием этой части
мозга. Гипоталамус имеет связи с конечным мозгом. Сам конечный мозг
разрастается и состоит из обонятельных луковиц и парных полушарий. В полушариях
у акул находятся зачатки старой коры (археокортекса) и древней коры
(палеокортекса).
Палеокортекс, тесно связанный с обонятельными луковицами,
служит главным образом для восприятия обонятельных стимулов. Археокортекс, или
гиппокампальная кора, предназначен для более сложной обработки обонятельной
информации. Вместе с тем электрофизиологические исследования показали, что
обонятельные проекции занимают только часть полушарий переднего мозга. Кроме
обонятельной здесь обнаружено представительство зрительной и соматической
сенсорных систем. Очевидно, старая и древняя кора может участвовать в регуляции
поисковых, пищевых, половых и оборонительных рефлексов у хрящевых рыб, многие
из которых являются активными хищниками.
Таким образом, у хрящевых рыб складываются основные черты
ихтиопсидного типа организации мозга. Его отличительной чертой является
присутствие надсегментарного аппарата интеграции, координирующего работу
моторных центров и организующего поведение. Эти интегративные функции
осуществляют средний мозг и мозжечок, что позволяет говорить о мезэнцефалоцеребеллярной
системе интеграции на данном этапе филогенетического развития нервной
системы. Конечный мозг участвует в регуляции функций нижележащих отделов.
Переход позвоночных от водного к наземному образу жизни
связан с целым рядом перестроек в ЦНС. Так, например, у амфибий в спинном мозгу
появляется два утолщения, соответствующих тазовому и грудному поясам
конечностей. В спинальных ганглиях вместо биполярных чувствительных нейронов
сосредоточиваются униполярные с Т—образно ветвящимся отростком, обеспечивающие
высокую скорость проведения возбуждения без участия клеточного тела. На
периферии в коже земноводных формируются специализированные рецепторы и рецепторные
поля, обеспечивающие дискриминационную чувствительность. В мозговом стволе
также происходят структурные изменения в связи с перераспределением
функциональной значимости различных отделов. В продолговатом мозгу наблюдаются
редукция ядер боковой линии и формирование улиткового слухового ядра,
осуществляющего анализ информации от примитивного органа слуха (см. разд. 4.6).
По сравнению с рыбами у амфибий, имеющих довольно
стереотипную локомоцию, наблюдается значительная редукция объема мозжечка.
Средний мозг, так же как и у рыб, представляет собой многослойную структуру, в
которой наряду с ростральными холмиками — ведущим отделом интеграции
зрительного анализатора — имеются дополнительные бугорки — предшественники
каудальных холмиков пластинки крыши.
Наиболее существенные в эволюционном плане изменения
происходят в промежуточном мозгу амфибий. Здесь обособляется таламус
(зрительный бугор), дифференцируются структурированные ядра (наружное
коленчатое тело) и восходящие пути, связывающие зрительный бугор с корой
(таламокортикальный путь).
В полушариях переднего мозга происходит дальнейшая
дифференциация зачатков старой и древней коры. В старой коре (археокортексе)
обнаруживаются звездчатые и пирамидные клетки. В промежутке между старой и
древней корой появляется полоска плаща, которая является предтечей новой
коры (неокортекса).
В целом развитие переднего мозга создает предпосылки для
перехода от свойственной рыбам мезэнцефалоцеребреллярной системы интеграции к диэнцефалотелэнцефалъной,
где ведущим отделом становится передний мозг, а
таламус промежуточного мозга
превращается в коллектор всех афферентных сигналов. В полной мере эта система
интеграции представлена в зауропсидном типе мозга у рептилий и знаменует собой
следующий этап морфофункциональной эволюции мозга.
Развитие таламокортикальной системы связей у рептилий
приводит к формированию новых проводящих путей, как бы подтягивающихся к
филогенетически молодым формациям мозга.
В боковых столбах спинного мозга рептилий обособляется
восходящий спинно—таламический путь, который проводит к головному мозгу
информацию о температурной и болевой чувствительности. Здесь же в боковых
столбах формируется новый нисходящий путь — красно—ядерно—спинномозговой
(Монакова). Он связывает мотонейроны спинного мозга с красным ядром среднего
мозга, которое включено в древнюю экстрапирамидную систему двигательной
регуляции. Эта многозвенная система объединяет влияние переднего мозга,
мозжечка, ретикулярной формации ствола, ядер вестибулярного комплекса и
координирует двигательную активность. У рептилий, как истинно наземных
животных, возрастает роль зрительной и акустической информации, возникает
необходимость сопоставления этой информации с обонятельной и вкусовой. В
соответствии с этими биологическими изменениями в стволовой части мозга рептилий
происходит целый ряд структурных изменений. В продолговатом мозгу
дифференцируются слуховые ядра, помимо улиткового ядра появляется угловое,
связанное со средним мозгом. В среднем мозгу двухолмие преобразуется в
четверохолмие, в ростральных холмах которого находятся акустические центры.
Наблюдается дальнейшая дифференциация связей крыши среднего
мозга с таламусом, который является как бы преддверия входа в кору всех
восходящих сенсорных путей. В самом таламусе происходит дальнейшее обособление
ядерных структур и установление между ними специализированных связей.
|
|
|
Рис. 3.1 Головной
мозг позвоночных животных А — рыба, Б — лягушка, В —
птица, Г — человек. Римскими
цифрами обозначены черепные нервы. |
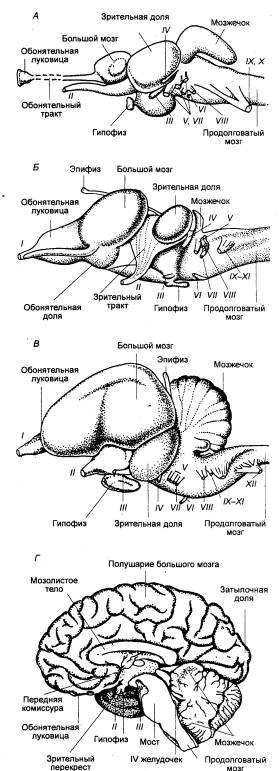
Конечный мозг рептилий может иметь два типа организации:
кортикальный и стриатальный. Кортикальный тип организации, свойственный
современным черепахам, характеризуется преимущественным развитием полушарий
переднего мозга и параллельным развитием новых отделов мозжечка. В дальнейшем
это направление эволюции мозга сохраняется у млекопитающих.
Стриатальный тип организации, характерный для
современных ящериц, отличается доминирующим развитием находящихся в глубине
полушарий базальных ядер, в частности полосатого тела. По этому пути далее идет
развитие головного мозга у птиц. Представляет интерес, что в полосатом теле у
птиц имеются клеточные объединения, или ассоциации нейронов (от трех до
десяти), разделенные олигодендроглией. Нейроны таких ассоциаций получают
одинаковую афферентацию, и это делает их сходными с нейронами, объединенными в
вертикальные колонки в новой коре млекопитающих. В то же время в полосатом теле
млекопитающих идентичные ассоциации не описаны. Очевидно, это является примером
конвергентной эволюции, когда сходные образования развились независимо у
различных животных.
У млекопитающих развитие переднего
мозга сопровождалось бурным ростом новой коры, находящейся в тесной
функциональной связи с таламусом промежуточного мозга (см. рис. 3.1). В коре
закладываются эфферентные пирамидные клетки, посылающие свои длинные аксоны к
мотонейронам спинного мозга.
Таким образом, наряду с многозвенной
экстрапирамидной системой появляются прямые пирамидные пути, которые
обеспечивают непосредственный контроль над двигательными актами. Корковая
регуляция движений у млекопитающих приводит к развитию филогенетически наиболее
молодой части мозжечка — передней части задних долей полушарий, или неоцеребеллума.
Неоцеребеллум приобретает двусторонние связи с новой корой.
Рост новой коры у млекопитающих
происходит настолько интенсивно, что старая и древняя кора оттесняются в
медиальном направлении к мозговой перегородке. Бурный рост коры компенсируется
формированием складчатости. У наиболее низко организованных однопроходных
(утконос) на поверхности полушария закладываются первые две постоянные борозды,
остальная же поверхность остается гладкой (лиссэнцефалический тип коры).
Как показали нейрофизиологические
исследования, мозг однопроходных и сумчатых млекопитающих лишен еще соединяющего
полушария мозолистого тела и характеризуется перекрытием сенсорных проекций в
новой коре. Четкая локализация моторных, зрительных и слуховых проекций здесь
отсутствует.
У плацентарных, млекопитающих
(насекомоядных и грызунов) отмечается развитие более четкой локализации
проекционных зон в коре. Наряду с проекционными зонами в новой коре формируются
ассоциативные зоны, однако границы первых и вторых могут перекрываться. Мозг
насекомоядных и грызунов характеризуется наличием мозолистого тела и дальнейшим
увеличением общей площади новой коры, развитием борозд и извилин (гирэнцефалический
тип коры).
В процессе параллельно—адаптивной эволюции у хищных
млекопитающих появляются теменные и лобные ассоциативные поля,
ответственные за оценку биологически значимой информации, мотивацию поведения и
программирование сложных поведенческих актов. Наблюдается дальнейшее развитие
складчатости новой коры.
И наконец, приматы демонстрируют наиболее высокий уровень
организации коры головного мозга. Кора приматов характеризуется
шестислойностью, отсутствием перекрытия ассоциативных и проекционных зон. У
приматов формируются связи между фронтальными и теменными ассоциативными полями
и, таким образом, возникает целостная интегративная система больших полушарий.
В целом, прослеживая основные этапы эволюции мозга
позвоночных, следует отметить, что его развитие не сводилось просто к линейному
увеличению размеров. В различных эволюционных линиях позвоночных могли иметь
место независимые процессы увеличения размеров и усложнения цитоархитектоники
различных отделов мозга. Примером тому может служить сравнение стриатального и
кортикального типов организации переднего мозга позвоночных.
В процессе развития наблюдается тенденция к перемещению
ведущих интегративных центров мозга в ростральном направлении от среднего мозга
и мозжечка к переднему мозгу, но эту тенденцию нельзя абсолютизировать, так как
мозг — это целостная система, в которой стволовые части играют важную
функциональную роль на всех этапах филогенетического развития позвоночных.
Кроме того, начиная с круглоротых, в переднем мозгу обнаруживаются проекции
различных сенсорных модальностей, что говорит об участии этого отдела мозга в
управлении поведением уже на ранних стадиях эволюции позвоночных.
3.2. ОБЩИЕ ПРИНЦИПЫ КООРДИНАЦИОННОЙ
ДЕЯТЕЛЬНОСТИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Интегративная деятельность ЦНС
сводится к соподчинению и объединению всех функциональных элементов организма в
целостную систему, обладающую определенной направленностью действия. В
осуществлении интегративной функции принимают участие различные уровни
организации ЦНС.
3.2.1. Интегративная и координационная деятельность нервной
клетки
Элементарной единицей ЦНС является нейрон (нервная
клетка, нейроцит), клеточная мембрана которого представляет поле, на котором
происходит интеграция синоптических влияний. Этот первый уровень
интеграции осуществляется в результате взаимодействия возбуждающих (ВПСП) и
тормозных (ТПСП) постсинаптических потенциалов, которые генерируются при
активации синаптических входов нейрона. В том случае, если возбуждающие и
тормозные входы активируются одновременно, происходит суммация синаптических
потенциалов противоположной полярности и мембранный потенциал в меньшей степени
приближается к критическому уровню деполяризации (КУД), при котором в
низкопороговой зоне клетки возникает потенциал действия. В некоторых случаях
снижение амплитуды ВПСП может происходить без возникновения ТПСП, только за
счет шунтирующего закорачивающего действия мембранных каналов, ответственных за
генерацию ТПСП.
Таким образом, конвергенция возбуждающих и тормозных входов
на мембране нейрона определяет частоту генерируемых им импульсных разрядов и
выступает в качестве универсального фактора интегративной деятельности нервной
клетки (см. разд. 1.1.8).
Координационная деятельность нейронов и слагаемых из них
элементарных нервных сетей (второй уровень интеграции) обусловлена
спецификой морфологических отношений в ЦНС. Довольно часто наблюдается такое
явление, когда одно пресинаптическое волокно многократно ветвится и образует
синаптические контакты сразу со многими нейронами (рис. 3.2). Это явление
|
|
|
Рис. 3.2 Дивергенция (А), конвергенция (Б), окклюзия (В),
временная (Г) и пространственная (Д) суммация в локальных нейронных цепях
(объяснение см. в тексте) |
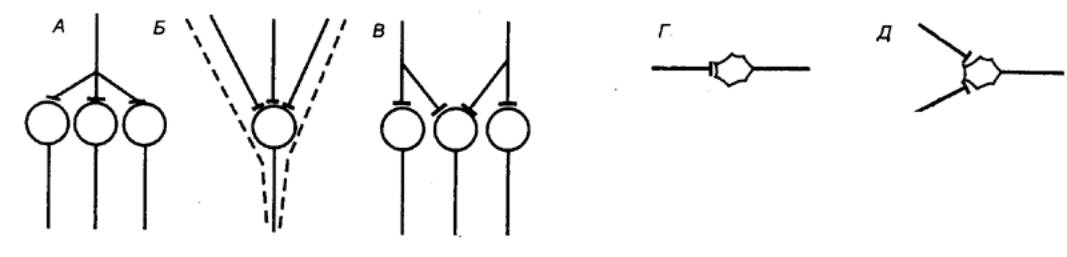
называется дивергенцией и
обнаруживается практически во всех отделах ЦНС: в организации афферентного
входа спинного мозга, в вегетативных ганглиях, в головном мозгу. Функционально
принцип дивергенции лежит в основе иррадиации возбуждения в рефлекторных
дугах, проявляющейся в том, что раздражение одного афферентного волокна может
вызвать генерализованную реакцию за счет возбуждения многих вставочных и
моторных нейронов.
3.2.2. Принцип общего конечного
пути
В структурной организации нервных
сетей встречается такая ситуация, когда на одном нейроне сходятся несколько
афферентных терминалей из других отделов ЦНС (рис. 3.2). Это явление принято
называть конвергенцией в нейронных связях. Так, например, к одному
мотонейрону подходит около 6000 коллатералей аксонов первичных афферентов,
спинальных интернейронов, нисходящих путей из стволовой части мозга и коры. Все
эти терминальные окончания образуют на мотонейроне возбуждающие и тормозные
синапсы и формируют своеобразную «воронку», суженная часть которой представляет
общий моторный выход. Данная воронка является анатомическим
образованием, определяющим один из механизмов координационной функции спинного
мозга (рис. 3.3).
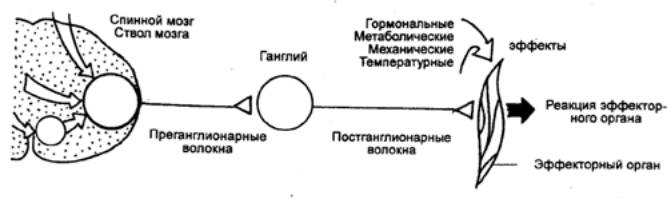
Суть этого механизма была раскрыта
английским физиологом Ч. Шеррингтоном, который сформулировал принцип общего
конечного пути. По Ч. Шеррингтону, количественное преобладание
чувствительных и других приходящих волокон над двигательными создает неизбежное
столкновение импульсов в общем конечном пути, которым являются группа
мотонейронов и иннервируемые ими мышцы. В результате этого столкновения
достигается торможение всех возможных степеней свободы двигательного аппарата,
кроме одной, в направлении которой протекает рефлекторная реакция, вызванная
максимальной стимуляцией одного из афферентных входов.
Рассмотрим случай с одновременным
раздражением рецептивных полей чесательного и сгибательного рефлексов, которые
реализуются идентичными группами мышц. Импульсы, идущие от этих рецептивных
полей, приходят к одной и той же группе мотонейронов, и здесь в узком месте
воронки за счет интеграции синаптических влияний осуществляется выбор в пользу
сгибательного рефлекса, вызванного более сильным болевым раздражением. Принцип
общего конечного пути, как один из принципов координации, действителен не
только для спинного мозга, он применим для любого этажа ЦНС, в том числе для
моторной коры.
|
|
|
Рис. 3.3 Нейронный
субстрат общего конечного пути автономной нервной системы |
3.2.3. Временная и пространственная суммация. Окклюзия
Конвергенция лежит в основе таких физиологических феноменов,
как временная и пространственная суммация. В том случае, если два
подпороговых раздражителя, приходящие к нейрону через афферентный вход (см.
рис. 3.2), следуют друг за другом с малым временным интервалом, имеет место
суммация вызванных этими раздражителями ВПСП, и суммарный ВПСП достигает
порогового уровня, достаточного для генерации импульсной активности. Данный
процесс способствует усилению поступающих к нейрону слабых сигналов и
определяется как временная суммация. Вместе с тем синаптическая
активация нейрона может осуществляться через два раздельных входа,
конвергирующих на эту клетку. Одновременная стимуляция этих входов
подпороговыми раздражителями также может привести к суммации ВПСП, возникающих
в двух пространственно разделенных зонах клеточной мембраны. В данном случае
происходит пространственная суммация, которая, так же как и временная,
может вызывать длительную деполяризацию клеточной мембраны и генерацию ритмической
импульсной активности на фоне этой деполяризации.
Однако возможна и такая ситуация, когда при одновременной
стимуляции двух входов возбуждение нейрона и соответствующий ему рефлекторный
ответ будут меньше алгебраической суммы ответов при раздельной стимуляции этих
входов. Причину явления можно уяснить из рис. 3.2. При раздельной стимуляции
двух входов мотонейрон б будет возбуждаться дважды: сначала совместно с
нейроном а и затем совместно с нейроном б. При одновременной стимуляции
двух входов нейрон б будет возбуждаться только один раз и,
соответственно, рефлекторный ответ будет меньше алгебраической суммы ответов
при раздельной стимуляции. Этот физиологический феномен, связанный с наличием
дополнительного общего пути для двух входов, получил название окклюзии.
Как уже отмечалось (см. разд. 1.1.8), локальные нейронные
сети могут усиливать слабые сигналы по механизму положительной обратной связи
за счет циклической реверберации возбуждения в цепи нейронов. Другой
возможный механизм усиления создается за счет синоптической потенциации
(облегчения) при ритмических раздражениях пресинаптических входов. Потенциация
выражается в увеличении амплитуды ВПСП во время (тетаническая потенциация) и
после (посттетаническая потенциация) ритмического раздражения пресинаптического
аксона с довольно высокой частотой (100—200 имп/с).
Это явление имеет гомосинаптическую природу, т. е.
возникает в том случае, когда ритмическое раздражение и пробный импульс
(вызывающий ВПСП) поступают к нейрону по одним и тем же афферентным волокнам.
Синаптическая потенциация при ритмических раздражениях может быть обусловлена:
1) повышением потенциала покоя мембраны пресинаптического аксона, ростом
амплитуды потенциала действия и, как следствие, высвобождением большего
количества медиатора из пресинаптической терминали; 2) мобилизацией готового к
выделению медиатора; 3) возрастанием внутриклеточной концентрации ионов Са2+,
который входит в пресинаптическую терминаль во время потенциала действия.
Координирующая функция локальных нейронных сетей помимо
усиления может выражаться и в ослаблении слишком интенсивной активности
нейронов за счет их торможения. Торможение, как особый нервный процесс,
характеризуется отсутствием способности к активному распространению по нервной
клетке и может быть представлено двумя формами — первичным и вторичным
торможением.
|
|
|
Рис. 3.4
Реципрокное (А), пресинаптическое
(Б) и возвратное (В) торможение в локальных нейронных цепях спинного мозга 1 — мотонейрон, 2 —
тормозный интернейрон, 3 — афферентные терминали. |
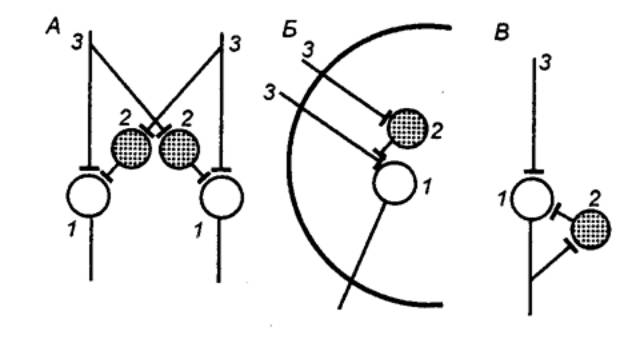
Первичное торможение обусловлено наличием специфических
тормозных структур и развивается первично без предварительного возбуждения.
Примером первичного торможения является так называемое реципрокное торможение
мыщц—антагонистов, обнаруженное в спинальных рефлекторных дугах (рис. 3.4).
Суть явления состоит в том, что если активируются проприорецепторы
мышцы—сгибателя, то они через первичные афференты одновременно возбуждают
мотонейрон данной мышцы—сгибателя и через коллатераль афферентного волокна —
тормозный вставочный нейрон. Возбуждение вставочного нейрона приводит к
постсинаптическому торможению мотонейрона антагонистической мышцы—разгибателя,
на теле которого аксон тормозного интернейрона формирует специализированные
тормозные синапсы. Реципрокное торможение играет важную роль в автоматической
координации двигательных актов.
Другим примером первичного торможения является открытое Б.
Реншоу возвратное торможение (рис. 3.4). Оно осуществляется в нейронной
цепи, которая состоит из мотонейрона и вставочного тормозного нейрона — клетки
Реншоу. Импульсы от возбужденного мотонейрона через отходящие от его аксона
возвратные коллатерали активируют клетку Реншоу, которая в свою очередь
вызывает торможение разрядов данного мотонейрона. Это торможение реализуется за
счет функции тормозных синапсов, которые клетка Реншоу образует на теле
активирующего ее мотонейрона. Таким образом, из двух нейронов формируется
контур с отрицательной обратной связью, позволяющий стабилизировать частоту
разрядов моторной клетки и подавить идущую к мышцам избыточную импульсацию.
В ряде случаев клетки Реншоу формируют тормозные синапсы не
только на активирующих их мотонейронах, но и на соседних мотонейронах со
сходными функциями. Осуществляемое через эту систему торможение окружающих
клеток называется латеральным.
Торможение по принципу отрицательной обратной связи
встречается не только на выходе, но и на входе моторных центров спинного мозга.
Явление подобного рода описано в моносинаптических соединениях афферентных
волокон со спинальными мотонейронами, торможение которых при данной ситуации не
связано с изменениями в постсинаптической мембране. Последнее обстоятельство
позволило определить эту форму торможения как пресинаптическое. Оно
обусловлено наличием вставочных тормозных нейронов, к которым подходят
коллатерали афферентных волокон (рис. 3.4). В свою очередь, вставочные нейроны
формируют аксо—аксональные синапсы на афферентных терминалях, являющихся
пресинаптическими по отношению к мотонейронам. В случае избыточного притока
сенсорной информации с периферии происходит активация тормозных интернейронов,
которые через аксо—аксональные синапсы вызывают деполяризацию афферентных
терминалей и, таким образом, уменьшают количество выделяемого из них медиатора,
а следовательно, и эффективность синаптической передачи. Электрофизиологическим
показателем этого процесса является снижение амплитуды регистрируемых от
мотонейрона ВПСП. Вместе с тем никаких признаков изменений ионной проницаемости
или генерации ТПСП в мотонейронах не наблюдается.
Вопрос о механизмах пресинаптического торможения
является довольно сложным. По—видимому, медиатором в тормозном аксо—аксональном
синапсе является ГАМК, которая вызывает деполяризацию афферентных терминалей за
счет увеличения проницаемости их мембраны для ионов Сl— . Деполяризация снижает амплитуду
потенциалов действия в афферентных волокнах и тем самым уменьшает квантовый
выброс медиатора в синапсе. Другой возможной причиной деполяризации терминалей
может быть повышение наружной концентрации ионов K+ при
длительной активации афферентных входов. Следует отметить, что феномен
пресинаптического торможения обнаружен не только в спинном мозгу, но и в других
отделах ЦНС.
В рассказе о координирующей роли торможения в локальных
нейронных цепях, следует упомянуть еще об одной форме торможения — вторичном
торможении, которое возникает без участия специализированных тормозных
структур как следствие избыточной активации возбуждающих входов нейрона. Эту
форму торможения определяют как торможение Введенского (пессимум),
который открыл его в 1886 г. при исследовании нервно—мышечного препарата.
Торможение Введенского играет предохранительную роль и
возникает при чрезмерной активации центральных нейронов в полисинаптических
рефлекторных дугах. Оно выражается в стойкой деполяризации клеточной мембраны,
превышающей критический уровень и вызывающей инактивацию Na—каналов,
ответственных за генерацию потенциалов действия. Таким образом, процессы
торможения в локальных нейронных сетях уменьшают избыточную активность и
участвуют в поддержании оптимальных режимов импульсной активности нервных
клеток.
При объединении нескольких локальных сетей формируются
нервные центры, которые представляют собой комплекс элементов, необходимых и
достаточных для осуществления определенного рефлекса или более сложного
поведенческого акта. В свою очередь, нервные центры, расположенные в различных
отделах мозга, могут кооперироваться в так называемые распределенные системы,
которые координируют "деятельность организма в целом. Эти системы имеют
иерархическую структуру и представляют собой следующий, более высокий
интегративный уровень ЦНС, деятельность которого базируется на некоторых
общих принципах работы нервных центров. Одним из таких принципов является
открытый А. А. Ухтомским принцип доминанты.
Под доминантой А. А. Ухтомский понимал господствующий
очаг возбуждения, предопределяющий характер текущих реакций центров в данный
момент. Такой господствующий, или доминантный, центр может возникнуть в
различных этажах ЦНС при достаточно длительном действии гуморальных или
рефлекторных раздражителей и характеризуется: 1) наличием повышенной
возбудимости; 2) инерционностью, обусловленной длительными следовыми
процессами; 3) способностью к суммации и сопряженному торможению других
центров, функционально несовместимых с деятельностью центров доминантного
очага. В целом доминанта как состояние характеризуется своей направленностью и
создает определенный вектор поведения.
Указанные выше черты доминанты можно проследить на примере
обнимательного рефлекса у лягушек, который возникает в период спаривания в
результате гормональных воздействий. Легчайшее прикосновение к «брачным»
мозолям на больших пальцах передних конечностей самца тотчас вызывает
обнимательный рефлекс, что свидетельствует о наличии повышенной возбудимости
флексорных (сгибательных) центров конечностей. Раздражение кожи механическими,
химическими или электрическими стимулами приводит к усилению рефлекса, что
является показателем суммации посторонних раздражителей. И наконец, высокий
порог, необходимый для вызова защитных реакций, указывает на сопряженное
торможение других центров.
Рассматривая доминанту как общий принцип работы нервных
центров, А. А. Ухтомский считал, что она связана с возбуждением целого
созвездия, или констелляции нервных центров, которые временно
кооперируются при выполнении биологически важной функции. Это созвездие создает
динамический функциональный орган, сообщающий организму единство
действия в данный момент. Формирование констелляции нервных центров, согласно
А. А. Ухтомскому, может происходить за счет импульсных взаимодействий и
сонастраивания работающих центров на единый ритм активности (усвоение
ритма).
Сформулированный А. А. Ухтомским в 1923 г. принцип доминанты
не является архаизмом, так как современная физиология накопила много фактов,
свидетельствующих о правильности основных положений этой теории.
Сегодня не вызывает сомнения возможность повышения
возбудимости центров под влиянием рефлекторных и гуморальных воздействий. В
некоторых участках мозга (латеральный и передний гипоталамус, предоптическая
область) обнаружены центральные рецепторы, реагирующие на содержание в
крови питательных веществ, половых гормонов и других факторов. Возбуждение этих
рецепторов может стать источником так называемых мотиваций (пищевой,
половой и т. п.) и привести к формированию доминантных состояний
соответствующих нервных центров.
Согласно современным представлениям (функциональная система
Анохина), любой поведенческий акт, в том числе и условный рефлекс,
начинается с анализа и синтеза афферентной информации, которые включают в себя
доминирующее мотивационное возбуждение, устраняющее избыточные степени свободы.
Доминирующий очаг возбуждения может возникнуть не только за счет
гуморальных воздействий, но и под влиянием нервных сигналов, изменяющих
возбудимость центральных нейронов. Это хорошо иллюстрируется моделью корковой
доминанты путем поляризации анодом слабого постоянного тока сенсомоторной коры
кролика в месте представительства одной из конечностей. Нейроны, расположенные
в области такого искусственно созданного доминантного очага, обладают
способностью суммировать возбуждение, вызванное посторонним, ранее
индифферентным раздражителем, и увеличивать частоту своих импульсных разрядов в
ответ на световые и звуковые стимулы. В результате индифферентные раздражители
на фоне изменяющей возбудимость анодной поляризации начинают вызывать движения
соответствующей конечности. Таким образом, анодная поляризация, которая
является эквивалентом достаточно долгой нервной сигнализации, формирует
доминантное состояние нервного центра, которое может поддерживаться довольно
длительное время.
Эта инерционность доминанты, как уже отмечалось выше, обусловлена
длительными следовыми процессами, механизмы которых детально освещены
современной физиологией. В естественных условиях длительное следовое
возбуждение может быть обусловлено: 1) суммацией ВПСП, вызванных приходящими к
нейронам подпороговыми нервными импульсами; 2) синаптической потенциацией
(облегчением) при ритмическом раздражении пресинаптических входов; 3)
изменением концентрации ионов К+ в синаптической щели, который как
деполяризатор усиливает вхождение ионов Са2+ в пресинаптическое
окончание; 4) метаболическими следами, связанными с влиянием медиаторов на
циклазные системы постсинаптических клеток; 5) циклическими связями в ЦНС,
способными обеспечить следовую самостимуляцию центров.
Следует отметить, что способность к длительному хранению следовых
процессов выражена по—разному в различных отделах мозга. Так, например, если в
спинальных центрах постсинаптическая потенциация длится минуты, то в центрах
гиппокампа она сохраняется часы и даже сутки. Очевидно, такие
специализированные блоки памяти, как гиппокамп, могут быть не только
участниками доминантных констелляций, но и хранителями следовых процессов от
пережитых доминант, представляющих собой системную реакцию мозга.
Доминантное состояние как системная реакция наряду с
суммацией посторонних раздражении предполагает сопряженное торможение
конкурирующих центров. Феномен сопряженного торможения продемонстрирован во
многих отделах ЦНС, где оно связано с наличием коллатералей сенсорных нейронов
и специализированных вставочных нейронов. В качестве примера можно привести
сопряженное торможение между отдельными колонками (модулями) сенсо—моторной
коры (см. разд. 3.10.3), которое определяет степень вовлечения того или иного
структурного модуля в распределенную систему. Сама гипотеза колончатой организации
коры больших полушарий и объединения групп модулей была выдвинута В.
Маунткаслом (1957), который представлял головной мозг как распределенную
систему нейронных модулей, связанных между собой многократно и очень сложно.
Информация распространяется по такой системе по многим разным путям, и
доминирование того или иного из модулей составляет динамическое и изменчивое
свойство системы.
При всей привлекательности такого подхода к пониманию
функций мозга его нельзя признать оригинальным, так как еще в 1923 г. А. А.
Ухтомским было сформулировано представление о динамическом функциональном
органе как о временной кооперации нервных центров, сообщающей организму
единство действия.
Спинной мозг (рис. 3.5.) является филогенетически
наиболее древним отделом ЦНС. В примитивной форме у ланцетника он
представляет собой трубку, идущую вдоль всего тела. От спинного мозга отходят
вентральные (передние, или двигательные) и дорсальные (задние, или
чувствительные) корешки (рис. 3.6).
У ланцетника спинномозговых узлов (спинальных ганглиев) еще
нет, чувствительные клетки рассеяны по ходу нервов или лежат в дорсальных
отделах спинного мозга. У круглоротых уже намечается разделение спинного мозга
на серое вещество, содержащее клетки и составляющее центральную часть, и
окружающее его белое вещество из продольно идущих безмякотных волокон.
Появляются и спинномозговые узлы. Они лежат на дорсальных корешках и состоят из
биполярных чувствительных нейронов.
Более сложно устроен спинной мозг круглоротых. В связи с тем
что у них происходит миелинизация нервных волокон, серое и белое вещество
приобретают четкие границы. В сером веществе возникают вентральные и дорсальные
рога, в белом веществе — вентральные и латеральные канатики. Вентральный и
дорсальный корешки сливаются и дают начало смешанному нерву. Таким образом, спинной
мозг круглоротых является прототипом спинного мозга высших позвоночных.
С появлением конечностей у амфибий возникают шейное и
поясничное утолщения спинного мозга, в передних рогах серого вещества выделяются
медиальная и латеральная группы клеток, образуются нисходящие и восходящие
пути.
|
|
|
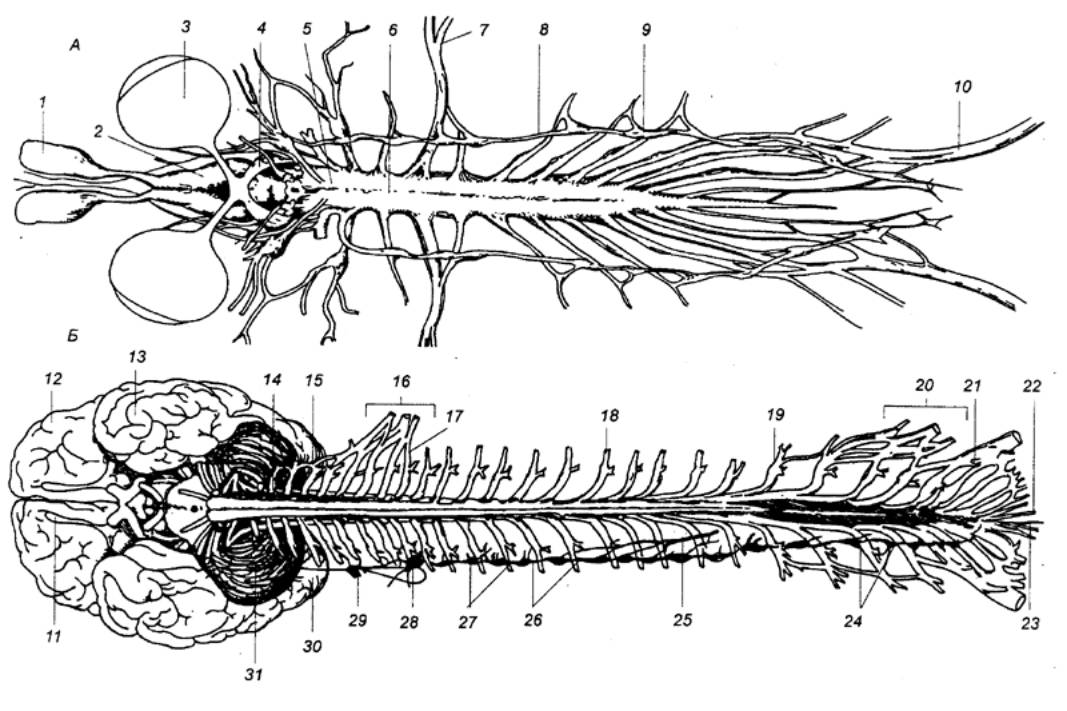
Рис. 3.5 Спинной
мозг лягушки (А) и человека (Б) с вентральной стороны 1 — носовой мешок, 2 — головной мозг, 3 — глаз, 4 — зрительный тракт, 5 —
продолговатый мозг, 6 —
спинной мозг, 7 — плечевой нерв, 8 — вегетативный ствол, 9 — спинальный ганглий, 10 — седалищный нерв, 11 — обонятельная луковица, 12 — лобная доля, 13 — височная доля, 14 — 1—й шейный нерв, 15 — мозжечок, 16 — плечевое сплетение, 17 — 1—й грудной нерв, 18 —спинальный ганглий, 19 — 1—й поясничный нерв, 20 — поясничное сплетение, 21 — 1—й крестцовый нерв, 22— копчиковый нерв, 23—концевая нить, 24 — соединительные ветви, 25 — чревный нерв, 26 — симпатический ствол, 27 — симпатический ганглий, 28 — нижний шейный симпатический
ганглий, 29 — средний шейный
симпатический ганглий, 30 —
затылочная доля, 31 — верхний
шейный симпатический ганглий. |
|
|
|
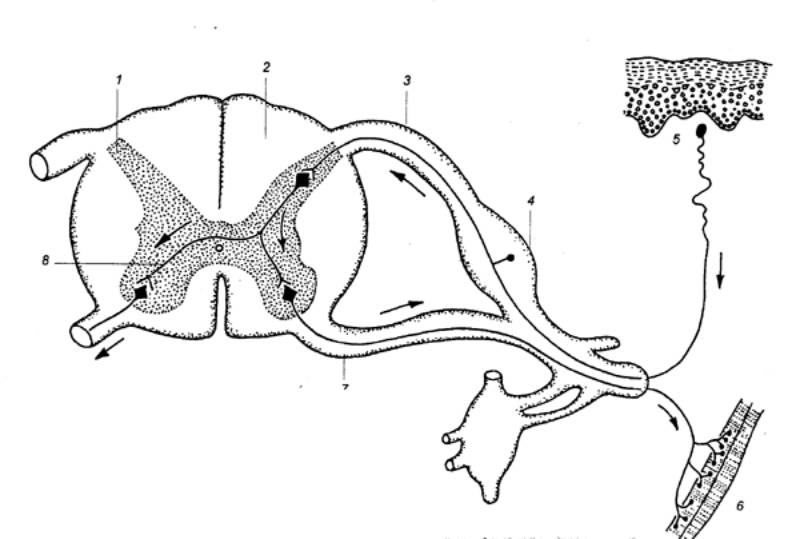
Рис. 3.6
Спинной мозг и его взаимосвязь с периферическими нервными волокнами— чувствительным,
двигательным. 1 — серое
вещество, 2 — белое вещество,
3 — дорсальный (задний) корешок, 4 —
спинальный ганглий, 5 — рецептор, 6
— эффектор, 7 —
вентральный (передний) корешок, 8 —
интернейрон. Стрелками показано направление распространения возбуждения. |
Дальнейшая дифференцировка
клеточно—волоконных структур происходит у рептилий и птиц. У них получают
развитие восходящие пути, лежащие в центральном и боковом канатиках, сформированы
дорсальный чувствительный путь и нисходящие связи. Происходит также
дифференциация ассоциативных клеток, возникновение межсегментарных
комиссуральных связей. У птиц хорошо развиты связи спинного мозга с
вестибулярным аппаратом и мозжечком.
Еще большая дифференцированность серого и белого вещества
наблюдается у млекопитающих. В дорсальных рогах появляются студенистое вещество
и грудное ядро, в вентральных — отчетливо выделяются клеточные группы,
возрастает число волокон в дорсальных и вентральных канатиках, появляется новый
восходящий путь от шейных сегментов к оливам.
В организации спинного мозга млекопитающих существуют
особенности, связанные с видовой принадлежностью животных. Спинной мозг у них
имеет разную длину, разное количество сегментов, неодинаковую выраженность
утолщений и пр. Все это зависит от числа позвонков, наличия хвоста, функции
конечностей.
При изучении деятельности спинного мозга необходимо
учитывать, что тело позвоночных животных и человека может быть разделено на пояса,
или сегменты. Сегменты, получающие чувствительные волокна от одной
отдельной пары дорсальных корешков, образуют метамер. Кожная область, в
которой распределяются эти чувствительные волокна, называется дерматомом
(рис. 3.7).
|
|
|
Рис. 3.7 Распределение дерматомов (областей чувствительной
иннервации) на поверхности тела Буквы соответствуют отделам спинного мозга: С — шейному, Т
— грудному, L — поясничному и S — крестцовому; цифрами показаны номера сегментов.
От каждого участка кожи, обозначенного одинаковыми символами, чувствительные
волокна отходят в один сегмент спинного мозга, например, чувствительность
кожи в области СЗ шеи обеспечивается 3—м шейным сегментом, область L5 на голени — 5—м поясничным
сегментом. |

|
|
|
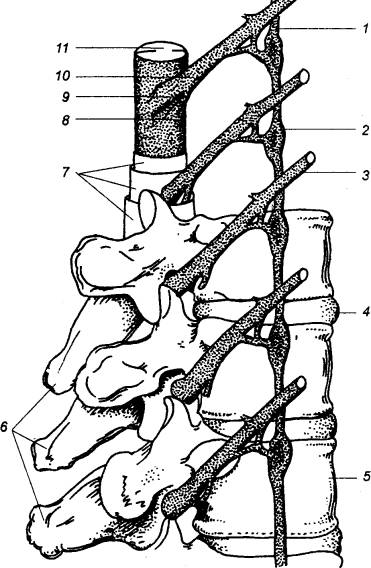
Рис. 3.8 Позвоночный столб, спинной мозг и пограничный
симпатический ствол 1 — симпатический ствол, 2 — симпатический ганглий, 3 —
спинномозговой нерв, 4 —
межпозвоночный диск, 5 —
позвонок, 6 — остистные
отростки позвонков, 7 — соединительнотканные оболочки, 8 — вентральный корешок, 9 — спинальный ганглий, 10 — дорсальный корешок, 11 — спинной мозг. |
В процессе эволюции тело позвоночных
изменило строение, в итоге возникли существенные отступления от идеальной метамерии.
Например, у ланцетника метамерия делит длину тела на неодинаковые отрезки.
Перестройку метамеров вызвало появление плавников у рыб. Метамерность особенно
усложнилась в связи с развитием конечностей и приобрела сложную форму для рук и
ног человека (рис. 3.7, 3.9).
Спинной мозг человека состоит из
следующих сегментов (обозначаются латинскими буквами): 8 шейных —C(I—8) 12 грудных — Т(1—12), 5 поясничных — L(1—5), 5 крестцовых —
S(1—5), 3 копчиковых — Со(1—3).
Общее количество сегментов соответствует числу метамеров тела, однако каждый
метамер получает иннервацию от двух—трех лежащих рядом сегментов. В сегментах
спинного мозга заканчиваются отростки подавляющего большинства чувствительных
нейронов тела, вступающие в составе дорсальных корешков. В спинном мозгу также
начинаются почти все эфферентные нервы организма: двигательные (за исключением
иннервирующих мышцы головы), все симпатические и часть парасимпатических. Они
следуют в составе дорсальных и вентральных корешков (рис. 3.8).
Если у лягушки перерезать справа
дорсальные, а слева вентральные корешки пояснично—крестцовых сегментов, то
правая лапка полностью теряет чувствительность, но может совершать движения.
Левая, наоборот, сохранит чувствительность, но окажется не способной к
двигательным реакциям. Это явление было открыто независимо друг от друга Ч.
Беллом (1811) и Ф. Мажанди (1822) и получило название закона Белла—Мажанди.
Характерной особенностью является
соотношение волокон, входящих через дорсальные и выходящих через вентральные
корешки. У кошки, например,
|
|
|
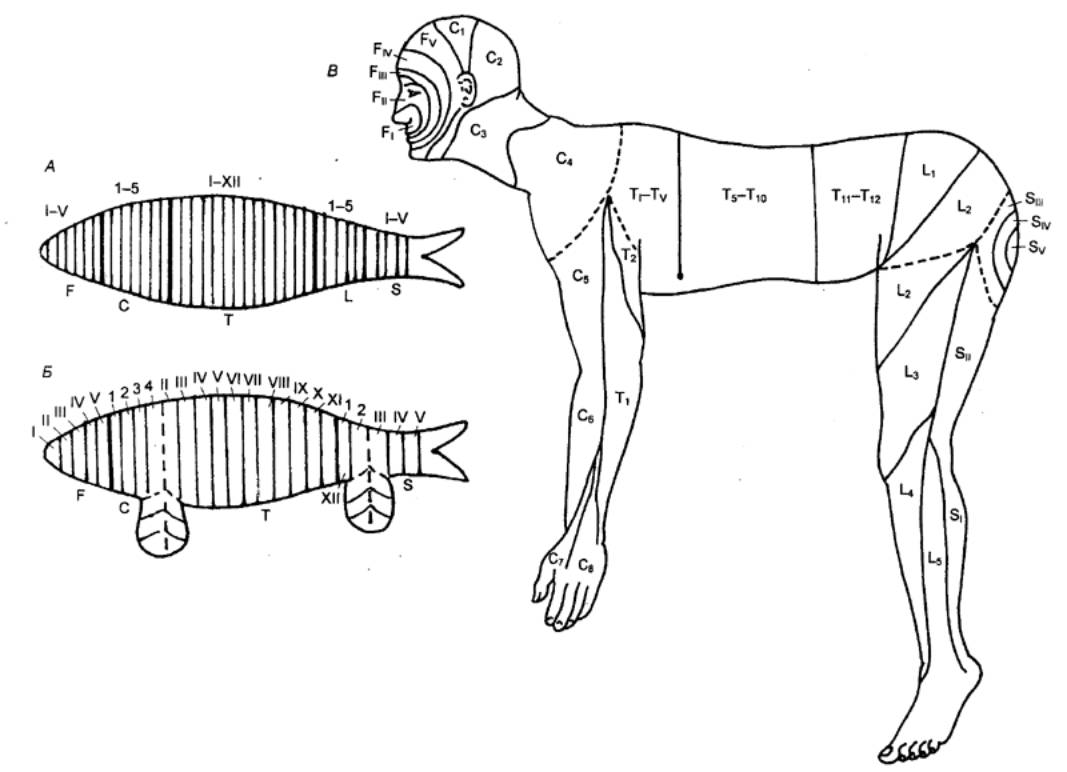
Рис.3.9 Схема
метамерности тела в процессе эволюции.
А— правильная
последовательность расположения метамеров (теоретически возможный случаи для
позвоночных); Б — нарушение правильной
метамерности тела рыб в связи с образованием плавников; В — то же у человека в связи с
образованием конечностей; F —
сегменты головы; С — сегменты шеи; Т — грудные сегменты; L — поясничные сегменты; S — крестцовые сегменты. |
дорсальный корешок поясничного
сегмента содержит около 12 000 волокон, тогда как вентральный — 6000.
Следовательно, один и тот же двигательный нейрон является общим конечным путем
для импульсов, поступивших от разных рецепторов и конкурирующих за общий конечный
путь. Несмотря на такую организацию, степень интегративной деятельности
спинного мозга ограничена по сравнению с другими вышележащими отделами ЦНС.
Классическими методами изучения функций спинного мозга
являются перерезки или разрушения его структур с последующей оценкой нарушения
функций. Большое развитие получило применение электрофизиологических методов,
включающих регистрацию суммарных электрических процессов, вне— и
внутриклеточные методы регистрации активности отдельных клеток.
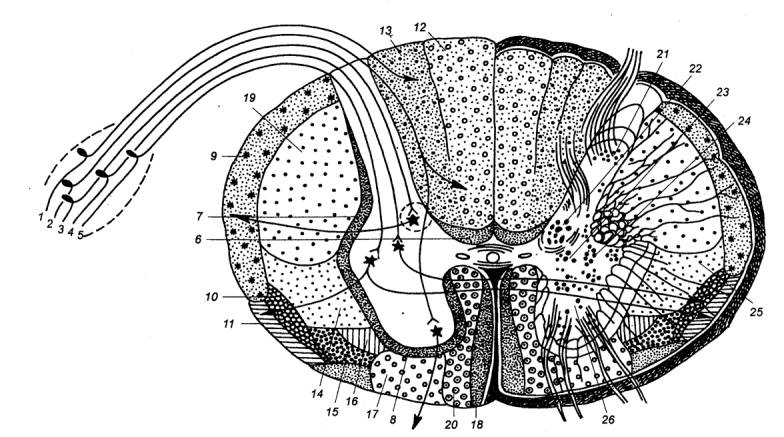
3.3.1. Нейронные структуры и их свойства
Тела чувствительных клеток вынесены за пределы спинного
мозга. Часть из них располагается в спинномозговых узлах (спинальных ганглиях)
(рис. 3.10, 3.11). Это тела соматических афферентов, иннервирующих в основном
скелетные мышцы. Другие находятся в экстра— и интрамуральных ганглиях
автономной нервной системы и обеспечивают чувствительность только внутренних
органов.
Чувствительные клетки имеют один отросток, который вскоре
после отхождения от тела клетки делится на две ветви (псевдоуниполярный
нейрон). Одна из них проводит возбуждение от рецепторов к телу клетки, другая —
от тела
|
|
|
Рис.3.10
Схема поперечного разреза спинного мозга и спинального ганглия.1,2 — рефлекторные пути сознательных
проприоцептивных ощущений и осязания, 3,4
— рефлекторные пути проприоцептивных импульсов, 5 — рефлекторные пути
температурной и болевой чувствительности, 6 — задний собственный пучок, 7 — боковой собственный пучок, 8 — передний собственный пучок, 9 — задний спинно—мозжечковый путь, 10 — передний спинно—мозжечковый путь, 11 — спинно—таламический путь, 12 — тонкий пучок, 13
— клиновидный пучок, 14 —
красно—ядерно—спинномозговой путь, 15
— таламо—спинномозговой путь, 16
— преддверно—спинномозговой путь, 17 — ретикулярно—спинномозговой путь, 18 — покрышечно—спинномозговой путь, 19 — латеральный корково—спинномозговой (пирамидный) путь, 20 — передний
корково—спинномозговой (пирамидный) путь, 21 — собственное ядро заднего рога, 22 — грудной столб (ядро), 23, 24 — промежуточное (серое) вещество, 25 — боковой промежуточный столб, 26 — ядра переднего рога. |
|
|
|
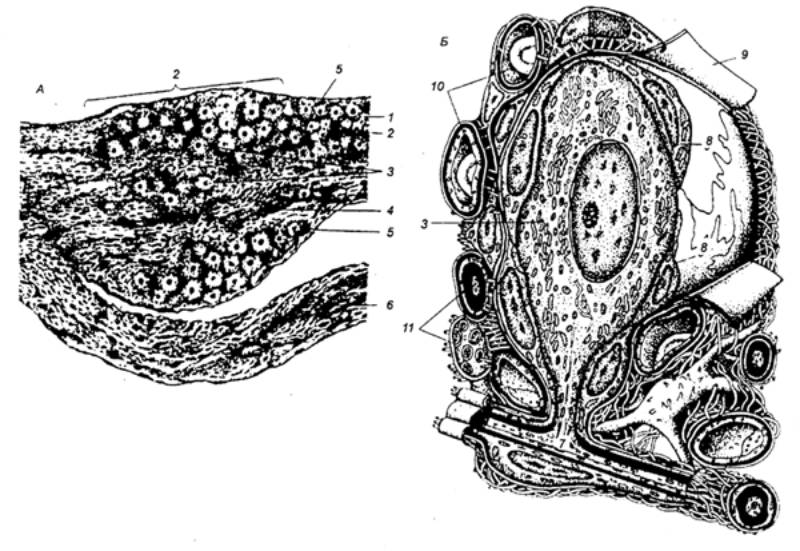
Рис. 3.11
Строение спинального ганглия.А —
продольный разрез ганглия и корешков; Б
— ультраструктурная организация псевдоуниполярного нейропита и его
микроокружения; 1 — дорсальный
корешок; 2 — ганглий; 3 —
псевдоуниполярные нейроциты; 4 —
нервные волокна; 5 — соединительнотканная капсула; 6 — вентральный корешок; 7 — Т—образное разветвление
отростка; 8 — мантийные
глиоциты; 9 — бааальная
мембрана; 10 — гемокапилляры; 11 —
миелиновые и безмиелиновые нервные волокна. |
|
|
|
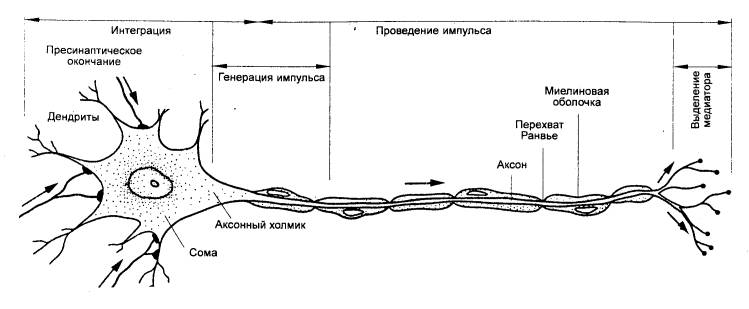
Рис. 3.12 Мотонейрон
спинного мозга позвоночных. Указаны функции разных его частей |
нервной клетки к нейронам спинного или
головного мозга. Распространение возбуждения из одной ветви в другую может
происходить без участия тела клетки.
Нервные волокна чувствительных
клеток по скорости проведения возбуждения и диаметру классифицируют на А—, В—
и С—группы. Толстые миелинизированные А—волокна с диаметром от 3 до 22
мкм и скоростью проведения возбуждения от 5 до 120 м/с разбиваются еще на
подгруппы: а — волокна от мышечных рецепторов, Р — от тактильных рецепторов и
барорецепторов, 8 — от терморецепторов, механорецепторов, болевых рецепторов. К
волокнам группы В относят миелиновые отростки средней толщины со
скоростью проведения возбуждения 3—14 м/с. По ним в основном передается
ощущение боли. К афферентным волокнам типа С относится большинство
безмиелиновых волокон толщиной не более 2 мкм и скоростью проведения 0,5—2 м/с.
Это волокна от болевых, хемо— и некоторых механорецепторов.
Сам спинной мозг в целом содержит, например, у человека
примерно 13 млн. нейронов. Из их общего числа только около 3% составляют
эфферентные, двигательные, или моторные, нейроны (рис. 3.12), а остальные 97%
приходятся на долю вставочных, или интернейронов. Двигательные нейроны являются
выходными клетками спинного мозга. Среди них различают α— и
γ—мотонейроны, a также преганглионарные нейроны автономной нервной системы.
α—Мотонейроны осуществляют передачу скелетным мышечным волокнам сигналов,
выработанных в спинном мозгу. Аксоны каждого мотонейрона многократно делятся,
и, таким образом, каждый из них охватывает своими терминалями до сотни мышечных
волокон, образуя в совокупности с ними двигательную единицу. В свою
очередь, несколько мотонейронов, иннервирующих ,одну и ту же мышцу, образуют мотонейронный
пул. В его состав могут входить мотонейроны нескольких соседних сегментов.
В связи с тем, что возбудимость / мотонейронов пула неодинакова, при слабых
раздражениях возбуждается только часть из них. Это влечет за собой
соответственно сокращение лишь части мышечных волокон. Другие моторные единицы,
для которых это раздражение является подпороговым, тоже реагируют, хотя их
реакция выражается лишь в деполяризации мембраны и повышении возбудимости. С
усилением раздражения они еще больше вовлекаются в реакцию, и, таким образом,
все двигательные единицы пула участвуют в рефлекторном ответе.
Максимальная частота воспроизведения ПД в α—мотонейроне
не превышает 200—300 имп/с. Вслед за ПД, амплитуда которого составляет 80—100
мВ, возникает следовая гиперполяризация длительностью от 50 до 150 мс.
По частоте импульсации и выраженности следовой гиперполяризации мотонейроны
разделяют на две группы: фазические и тонические. Особенности их возбуждения
коррелируют с функциональными свойствами иннервируемых мышц. Фазическими
мотонейронами иннервируются более быстрые, «белые» мышцы, тоническими —
более медленные, «красные» (см. разд. 1.2.1).
В организации функции α—мотонейронов важным звеном
является наличие системы отрицательной обратной связи, образованной
аксонными коллатералями и специальными тормозными вставочными нейронами — клетками
Реншоу. Своими возвратными тормозными влияниями они могут охватывать большие
группы мотонейронов, обеспечивая, таким образом, интеграцию процессов
возбуждения и торможения.
γ— Мотонейроны, иннервируют интрафузальные
(внутриверетенные) мышечные волокна. Они разряжаются с более низкой частотой, и
следовая гиперполяризация у них выражена слабее, чем у α—мотонейронов. Их
функциональное значение сводится к сокращению интрафузальных мышечных волокон,
не приводящему, однако, к появлению двигательного ответа. Возбуждение этих
волокон сопровождается изменением чувствительности их рецепторов к сокращению
или расслаблению экстрафузальных мышечных волокон.
Нейроны автономной нервной системы составляют особую группу клеток.
Тела симпатических нейронов, аксоны которых являются преганглионарными
волокнами, располагаются в боковом промежуточном столбе спинного мозга (см.
разд. 5.1.2). По своим свойствам они относятся к группе В—волокон. Характерной
особенностью их функционирования является низкая частота свойственной им
постоянной тонической импульсной активности. Одни из этих волокон участвуют в
поддержании сосудистого тонуса, другие — обеспечивают регуляцию висцеральных
эффекторных структур (гладкой мускулатуры пищеварительной системы, железистых
клеток).
Тела парасимпатических нейронов образуют крестцовые
парасимпатические ядра. Они располагаются в сером веществе крестцовых сегментов
спинного мозга. Для многих из них характерна фоновая импульсная активность,
частота которой возрастает по мере повышения давления в мочевом пузыре. При
раздражении висцеральных тазовых афферентных волокон в эфферентных клетках
регистрируется вызванный разряд, характеризующийся чрезвычайно большим
латентным периодом.
К вставочным, или интернейронам, спинного
мозга относятся нервные клетки, аксоны которых не выходят за его пределы. В
зависимости от хода отростков различают собственно спинальные и проекционные. Спинальные
интернейроны ветвятся в пределах нескольких смежных сегментов, образуя
внутрисегментарные и межсегментарные связи. Наряду с ними существуют
интернейроны, аксоны которых проходят через несколько сегментов или даже из
одного отдела спинного мозга в другой. Их аксоны образуют собственные пучки
спинного мозга.
К проекционным интернейронам
относятся клетки, длинные аксоны которых формируют восходящие пути спинного
мозга. На каждом интернейроне расположено в среднем около 500 синапсов.
Синаптические влияния в них опосредуются через ВПСП и ТПСП, суммация которых и
достижение критического уровня приводят к возникновению распространяющегося ПД.
Дорсальную часть дорсального рога спинного мозга занимает
характерное скопление нервных клеток, образующих густое сплетение. Эта область
получила название студенистого (роландова) вещества. У
позвоночных животных и человека послойное рассмотрение клеточной архитектуры
серого вещества спинного мозга показало существование последовательно
расположенных слоев, или пластин, состоящих из однотипных клеток.
Согласно Рекседу (Rexed), все серое вещество разделяется на 10 таких пластин (рис.
3.13).
|
|
|
Рис. 3.13 Послойная топография
серого вещества спинного мозга по Рекседу |
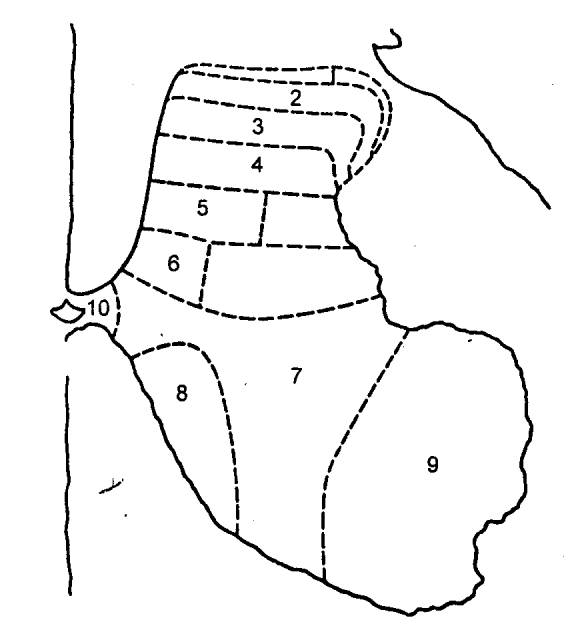
Из них первая — дорсальная, содержит преимущественно
нейроны, аксоны которых дают начало спинно—таламическому пути. На самих нейронах
оканчиваются первичных афференты и аксоны проприоспинальных нейронов. Вторая
и третья пластины образуют названное выше студенистое вещество. Здесь
выделяются
два вида нейронов с многочисленным
дендритным ветвлением — мелкие и крупные. Аксоны этих клеток в большинстве
оканчиваются здесь же в студенистом веществе, хотя некоторые из них
проецируются на дорсолатеральный собственный пучок спинного мозга. Четвертая
пластина локализуется в центре дорсального рога. Составляющие ее клетки
получают синаптический приток от нейронов студенистого тела и первичных
чувствительных нейронов, аксоны же проецируются в таламус и боковое шейное
ядро. Пятая и шестая пластины представлены конгломератом
разнообразных вставочных нейронов, получающих синаптический приток от волокон
дорсальных корешков, а также и от нисходящих путей — в основном
кортиково—спинномозгового и
красноядерно—спинномозгового путей. Седьмая и восьмая
пластины являются местом положения проприоспинальных вставочных нейронов с
длинными аксонами. На этих интернейронах оканчиваются аксоны проприорецепторов
и проприоспинальных нейронов, а также волокна преддверно—спинномозгового и
ретикулярно—спинномозгового путей. Девятая пластина является местом
сосредоточения α— и γ—мотонейронов. Этой зоны достигают пресинаптические
структуры первичных афферентов от мышечных рецепторов растяжения. Сюда приходят
также окончания волокон нисходящих трактов, аксоны возбуждающих и тормозящих
интернейронов. Наконец, десятая пластина. Она окружает спинномозговой
канал и характеризуется наряду с присутствием нервных клеток еще и наличием
значительного количества глиальных и комиссуральных волокон.
3.3.2. Рефлекторная функция спинного мозга
Важную роль в развитии представлений о рефлекторной
деятельности спинного мозга сыграли открытия и обобщения английского физиолога
Ч. Шеррингтона.
Объем функций, осуществляемых спинным мозгом, чрезвычайно
велик. В нем находятся центры: всех двигательных рефлексов (за исключением мышц
головы); всех рефлексов мочеполовой системы и прямой кишки; рефлексов,
обеспечивающих терморегуляцию; регулирующих метаболизм тканей; большинства
сосудистых рефлексов; сокращения диафрагмы и др. В естественных условиях эти
рефлексы всегда испытывают влияние высших отделов головного мозга.
Степень проявления рефлексов зависит от того, сохраняются ли
связи структур спинного мозга со структурами головного мозга. После децеребрации
(удаления головного мозга) или спинализации (отделения спинного мозга от
головного посредством перерезки) исчезают многие сложные формы активности,
создаваемые спинным мозгом. При этом определенное значение принадлежит уровню
организации подопытного животного. Например, спинальная лягушка, представитель
низших позвоночных, может сидеть и вырываться, когда ее схватывают. Спинальная
же собака сама не может ни стоять, ни ходить. Это объясняется тем, что
разобщение спинного мозга и вышележащих структур нарушает рефлекторные дуги,
ответственные за осуществление определенных реакций. При этом, в частности,
исчезают периодические разряды дыхательных мышц, обеспечивающие дыхательные
движения, пропадают тонические разряды симпатических нейронов, поддерживающих
сосудистый тонус и, соответственно, артериальное давление.
Как правило, рефлексы спинальных животных являются
координированными. Возбуждение у них каждой группы рецепторов сопровождается
своим специфическим ответом. Например, механическое раздражение кожи стопы у
лягушки вызывает сгибание раздражаемой конечности и разгибание другой.
Раздражение рецепторов мочевого пузыря и прямой кишки сопровождается рефлекторным
сокращением их мускулатуры.
В связи с отсутствием тонических влияний структур головного
мозга изменяется функциональное состояние и самих спинномозговых нейронных
систем. К числу таких нарушений относится прекращение сложных локомоторных
актов типа шагания. Характерно, что исчезновение этих актов после спинализации
удается восстановить введением животному веществ, способствующих выделению
медиаторов синаптическими окончаниями перерезанных путей.
|
|
|
Рис. 3.14 Внутрисегментарные связи сенсорного волокна от
болевого рецептора кожи стопы Р —
мотонейроны разгибателей; С — мотонейроны сгибателей. |
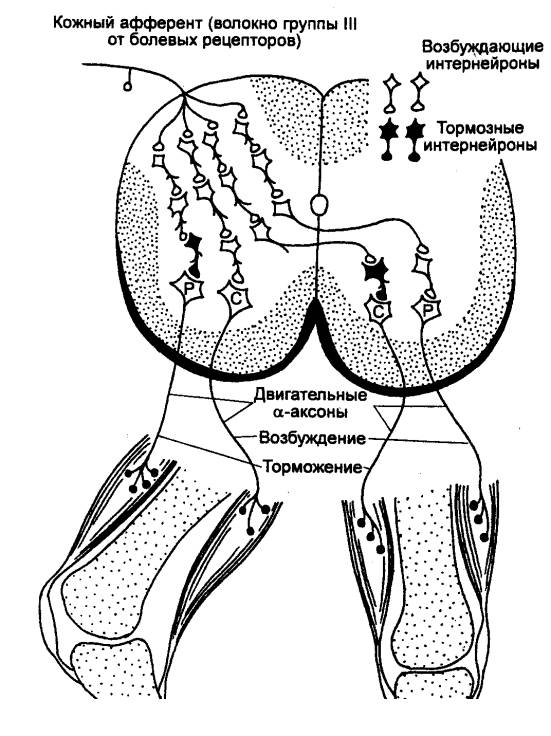
В зависимости от числа нейронов, участвующих в проведении возбуждения,
рефлекторные дуги спинного мозга делятся на моносинаптические и
полисинаптические.
Моносинаптическая дуга состоит из чувствительного нейрона
с рецепторами мышечных веретен и эффекторного нейрона, оканчивающегося на
мышечных волокнах. Классическим примером моносинаптической дуги может служить
рефлекторная дуга коленного рефлекса, в которой возбуждение от рецептора до
эффектора проходит всего за 0,5—1,0 мс, т. е. время, необходимое для
прохождения возбуждения всего лишь через один синапс.
В полисинаптической дуге на
пути возбуждения от рецептора к эффектору помимо чувствительного и эффекторного
нейрона находятся еще вставочные нейроны (рис. 3.14). Таким образом,
возбуждение в этой дуге проходит не через один, а через несколько синапсов,
определяющих время латентного периода ответа и суммарной синаптической
задержки. При осуществлении рефлекторных реакций
|
|
|
Рис. 3.15 Рефлексы, спинного мозга А — коленный; Б — ахиллов; В — подошвенный; Г — сгибательно—локтевой;
Д — разгибательно—локтевой; Е — брюшной. |
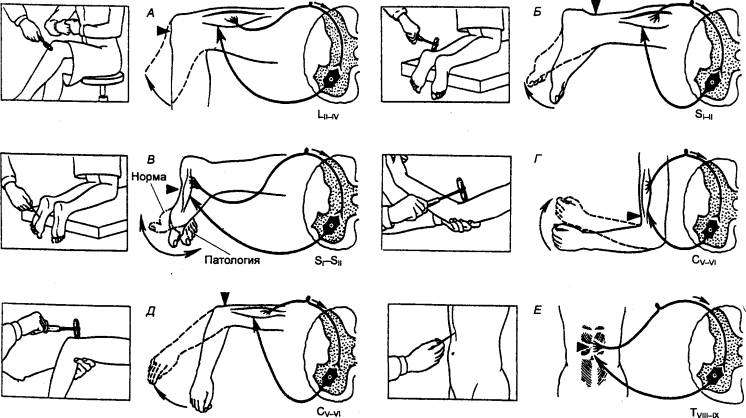
Таблица 3.1
Соматические спинномозговые рефлексы человека
|
Название
рефлекса |
Применяемое
раздражение |
Характер
рефлекторной реакции |
Локализация
нейронов, участвующих в рефлексе |
|
Сухожильные
проприорецептивные рефлексы: |
|
|
|
|
сгибательно—локтевой
(бицепс—рефлекс) |
Удар
молоточком по сухожилию двуглавой мышцы плеча (рука слегка согнута в локте) |
Сокращение
двуглавой мышцы плеча и сгибание руки |
V—VI шейные сегменты спинного мозга |
|
коленный |
Удар
молоточком по сухожилию четырехглавой мышцы бедра ниже надколенника |
Сокращение
четырехглавой мышцы бедра и разгибание голени |
II—IV поясничные сегменты |
|
ахиллов |
Удар по
ахиллову сухожилию |
Подошвенное
сгибание стопы |
1—11
крестцовые сегменты |
|
Брюшные
рефлексы: |
Штриховое
раздражение кожи: |
Сокращение
соответствующих участков брюшной мускулатуры |
|
|
верхний |
параллельно
нижним ребрам |
|
VIII—IX грудные сегменты |
|
средний |
на уровне
пупка (горизонтально) |
|
IX—XII грудные сегменты |
|
нижний |
параллельно
паховой складке |
|
I—II поясничные сегменты |
|
Кремастерный
яичковый рефлекс |
Штриховое
раздражение внутренней поверхности бедра |
Сокращение
мышцы, поднимающей яичко |
I—II поясничные сегменты |
|
Анальный
рефлекс |
Штрих
или укол вблизи заднего прохода |
Сокращение
наружного сфинктера прямой кишки |
IV—V крестцовые сегменты |
|
Подошвенный
рефлекс |
Слабое
штриховое раздражение подошвы |
Сгибание
пальцев и стопы |
I—II крестцовые сегменты |
|
Сильное
раздражение подошвы |
Разгибание
пальцев и сгибание ноги |
в моно— и полисинаптических дугах
вовлекаются также координирующие рефлекс многочисленные интернейроны. Подобный анализ
периферических воздействий, а также надсегментарный контроль за рефлекторной
деятельностью спинного мозга происходит с помощью длинных многоэтажных
рефлекторных дуг. Их центры локализованы в подкорковых областях и коре
головного мозга.
К числу рефлексов спинного мозга (рис. 3.15, табл.
3.1) относятся защитные рефлексы, рефлексы на растяжение, мышц—антагонистов,
висцеромоторные, вегетативные рефлексы. Эта классификация весьма условна, и
весь ее смысл в том, что она указывает на многообразие рефлекторных ответов.
Даже у спинального животного трудно встретить рефлексы, которые относились бы
только к одной из названных групп.
Раздражение в виде укола кожи стопы вызывает у лягушки защитный
рефлекс — либо отдергивание лапки при легком уколе, либо вовлечение в
реакцию другой лапки и отодвигание от источника при более сильном воздействии,
наконец, убегание животного при значительном болевом раздражении, когда в
возбуждение вовлекаются многие структуры нервной системы.
Рефлексы растяжения проявляются укорочением мышцы в
ответ на ее растяжение (рис. 3.16). Основными рецепторами в этом случае служат
нервно—мышечные веретена, афферентным звеном — чувствительные волокна
соматических нервов и дорсальных корешков спинного мозга. Эти рефлекторные дуги
чаще всего замыкаются в спинном мозге. Начало и конец рефлекторной дуги связаны
с мышцей. Рефлексы наиболее выражены в мышцах—разгибателях. Для того чтобы
организм мог противостоять силе земного притяжения, эти мышцы должны находиться
в состоянии тонического напряжения. Биологическое значение этих рефлексов
состоит в том, что они участвуют в сохранении статики и положения тела,
регулируя степень сокращения мышцы в соответствии с действующими на нее
раздражениями. Особое значение этот вид рефлексов имеет у копытных животных,
хотя и у человека они хорошо развиты.
|
|
|
Рис. 3.16
Дуга рефлекса растяжения и реципрокного торможения мышц—антагонистов С —
мотонейроны сгибателей коленного сустава; Р — мотонейроны разгибателей
коленного сустава. 1 — волокна Iа, 2 — двигательные
аксоны, 3 — концевые пластинки, 4 — сгибатель, 5 — разгибатель. |
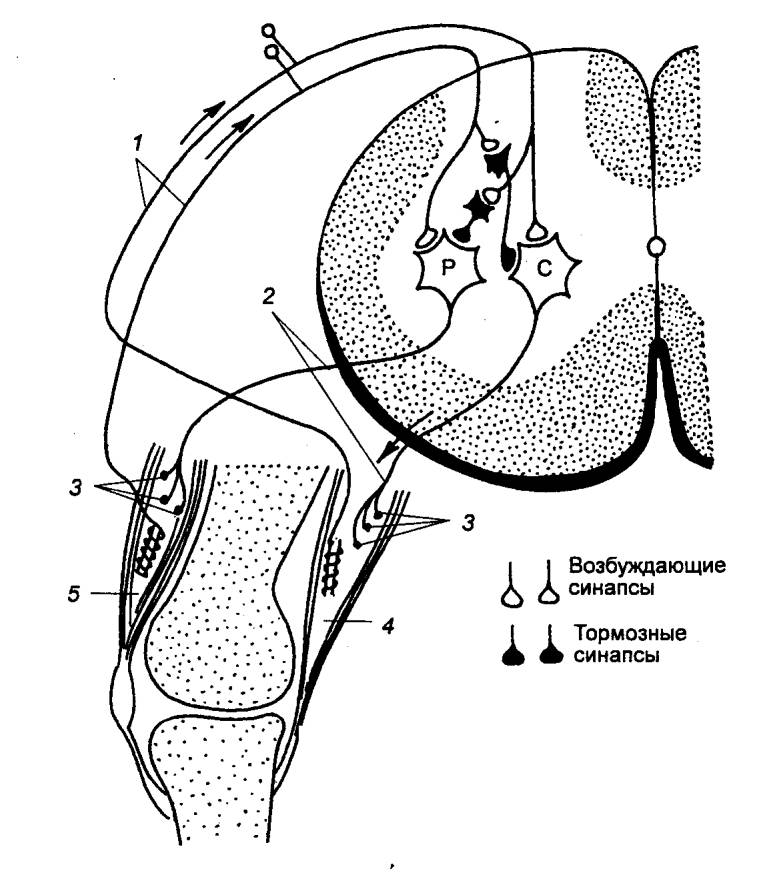
Рефлексы мышц—антагонистов (рис. 3.17) лежат в основе
локомоторных актов и характеризуются тем, что при возбуждении мотонейронов сгибателей
одновременно происходит торможение мотонейронов мыщц—разгибателей. При этом в
конечности другой стороны наблюдаются обратные явления. В целом это создает
правильное чередование противоположных по функциональному значению мышечных
сокращений. Механизм, обусловливающий такое чередование активности различных
двигательных ядер, например при ходьбе, локализуется в интернейронном аппарате
спинного мозга. Вместе с тем для его активации необходимо поступление
тонического нисходящего сигнала из двигательных центров головного мозга.
Висцеромоторные рефлексы возникают при возбуждении
афферентных волокон внутренних органов и характеризуются появлением
двигательных реакций мышц грудной и брюшной стенки, мышц—разгибателей спины.
Возникновение этих рефлексов связано с существованием конвергенции висцеральных
и соматических афферентных волокон к одним и тем же интернейронам спинного
мозга.
Вегетативные рефлексы заключаются, во—первых, в появлении
полисинаптических разрядов в преганглионарных симпатических волокнах в ответ на
возбуждение
|
|
|
Рис. 3.17 Нервные пути сгибательного и разгибательного рефлекса С —
ипсилатеральные сгибательные мотонейроны; Р — ипсилатеральные разгибательные
мотонейроны. 1 — белое
вещество, 2 — разгибатель, 3
— сгибатель. |
|
|
|
Рис. 3.18 Соединение кожных и висцеральных афферентное в спинном мозге I — кожа, 2 — механорецепторы, 3 — нисходящие пути, 4 — тормозный нейрон, 5 —
дорсальный столб, 6 — поясничный
отдел, 7 — грудной отдел, 8 — шейный
отдел, 9 — вентролатеральный
канатик, 10 — дорсальный
корешок, II —
вентральный корешок, 12 —
проприоспинальный путь, 13 —
двигательные аксоны, 14 —
симпатические аксоны, 15 —
вентральный рог, 16 — внутренности,
17 — висперопепторы, 18 — мышца, 19 — проприопепторы, 20
— дорсальный рог, 21 —
терморецепторы, 22 —
ноцицепторы. |
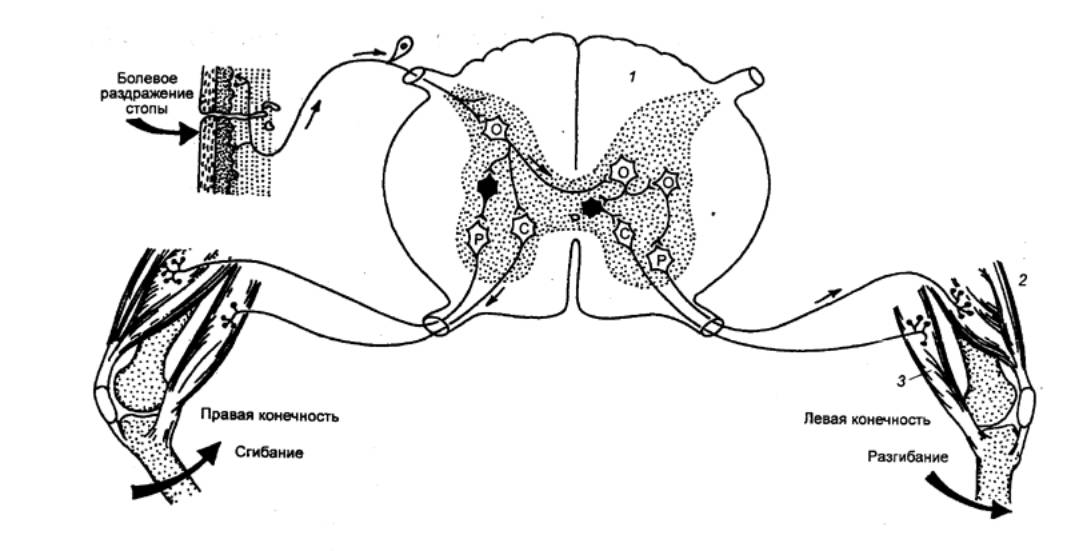
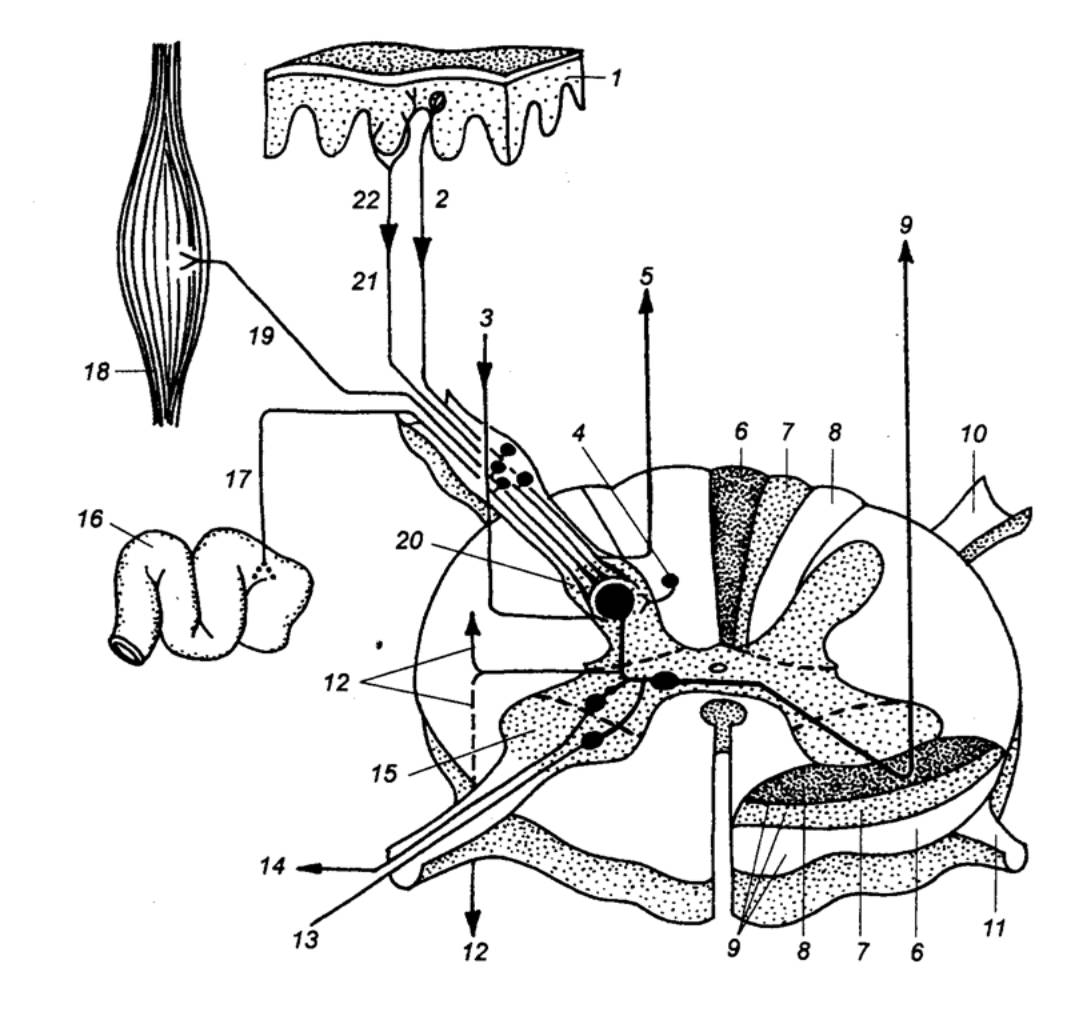
симпатических и соматических
чувствительных клеток (рис. 3.18), во—вторых, в возникновении рефлекторных
реакций парасимпатических нейронов в ответ на раздражение чувствительных путей (более
подробно эти реакции рассмотрены в разд. 5.3).
Наряду с выполнением собственных рефлекторных реакций
нейронные структуры спинного мозга служат аппаратом для реализации большого
числа сложных процессов, осуществляемых различными отделами головного мозга.
Это управление может быть прямым, когда нисходящие пути непосредственно связаны
с мотонейронами спинного мозга, и непрямым через интернейроны образующие
короткие межсегментарные связи. Свойства последних и особенности связей с
нисходящими волокнами и моторными нейронами создают возможность к интеграции
поступающих сигналов, их переработке и пространственному перераспределению.
3.3.3. Проводниковые функции спинного мозга
Помимо рефлекторной деятельности еще одной важной функцией
спинного мозга является проведение импульсов. Оно осуществляется белым
веществом, состоящим из нервных волокон.
В результате эволюционного развития простая рефлекторная
дуга, лежащая в основе функции нервной системы, усложняется, и в каждой ее
части вместо одного нейрона возникают цепи нервных клеток, аксоны которых
образуют проводящие пути. Под проводящими путями принято понимать группы
Таблица 3.2
Основные проводящие пути спинного мозга
|
Проводящие
пути |
Столбы
спинного мозга |
Физиологическое
значение |
|
А.
Восходящие (чувствительные) пути |
||
|
1.
Тонкий пучок (пучок Голля) |
Дорсальные |
Тактильная
чувствительность, чувства положения тела, пассивных движений тела, |
|
2.Клиновидный
(пучок Бурдаха) |
« |
Тоже |
|
3.
Дорсолатеральный |
Боковые |
Пути
болевой и температурной чувствительности |
|
4.
Дорсальный спинно—мозжечковый путь (пучок Флексига) |
« |
Импульсы
от проприоцепторов мышц, сухожилий, связок; чувство давления и прикосновения
из кожи |
|
5.
Вентральный спинно—мозжечковый путь (пучок Говерса) |
« |
Тоже |
|
таламический
путь |
« |
Болевая
и температурная чувствительность |
|
7.
Спинно—тектальный путь таламический путь |
« |
Сенсорные
пути зрительно—двигательных рефлексов (?) и болевой чувствительности (?) |
|
Вентральные |
Тактильная
чувствительность |
|
|
|
||
|
1.
Латеральный корково—спинномозговой (пирамидный) путь |
Боковые |
Импульсы
к скелетным мышцам. Произвольные движения |
|
2.
Красноядерно—спинномозговой (Монакова) путь |
« |
Импульсы,
поддерживающие тонус скелетных мышц |
|
спинномозговой
путь |
« |
позы и
равновесия тела |
|
4.
Оливоспинномозговой (Гельвёга) путь ' |
« |
Функция неизвестна.
Возможно, он участвует в осуществлении таламоспинальных рефлексов |
|
5.
Ретикулярно—спинномозговой путь |
Вентральные |
Импульсы,
поддерживающие тонус скелетных мышц, регулирующие состояние спинальных
вегетативных центров и чувствительность мышечных веретен проприорецепторов
скелетных мышц |
|
6.
Вентральный преддверно—спинномозговой путь |
« |
Импульсы,
обеспечивающие поддержание позы и равновесия тела |
|
7.
Тектоспинальный (покрышечно—спинномозговой) путь |
« |
Импульсы,
обеспечивающие осуществление зрительных и слуховых двигательных рефлексов |
|
8.
Вентральный корково—спинномозговой (пирамидный) путь |
Вентральные |
Импульсы
к скелетным мышцам, произвольные движения |
нервных волокон, характеризующиеся общностью
строения и функций. Они связывают различные отделы спинного мозга или спинной и
головной мозг (табл. 3.2). Все нервные волокна одного пути начинаются от
однородных нейронов и заканчиваются на нейронах, выполняющих одинаковую
функцию.
В соответствии с функциональными особенностями различают
ассоциативные, комиссуральные и проекционные (афферентные и эфферентные)
нервные волокна. Ассоциативные волокна, или пучки осуществляют
односторонние связи между отдельными частями спинного мозга. Связывая разные
сегменты, они образуют собственные пучки, являющиеся частью сегментарного
аппарата спинного мозга. Комиссуральные волокна соединяют функционально
однородные противоположные участки разных отделов спинного мозга. Проекционные
волокна связывают спинной мозг с вышележащими отделами. Эти волокна
образуют основные проводящие пути, которые представлены восходящими
(центростремительными, афферентными, чувствительными) и нисходящими
(центробежными, эфферентными, двигательными) путями.
Восходящие проводящие пути. Несут импульсы от рецепторов,
воспринимающих информацию из внешнего мира и внутренней среды организма. В
зависимости от вида чувствительности, которую они проводят, их делят на пути экстеро—,
проприо— и интероцептивной чувствительности. Нисходящие пути передают
импульсы от структур головного мозга к двигательным ядрам, осуществляющим
ответные реакции на внешние и внутренние раздражения.
Основными восходящими путями спинного мозга являются тонкий
пучок, клиновидный пучок, латеральный и вентральный спинно—таламические пути,
дорсальный и вентральный спинно—мозжечковые пути (рис. 3.19).
Тонкий пучок (Голля) и клиновидный пучок (Бурдаха) составляют
задние канатики спинного мозга. Эти пучки волокон являются отростками
чувствительных клеток спинальных ганглиев, проводящих возбуждение от
проприорецепторов мышц, сухожилий, частично тактильных рецепторов кожи,
висцерорецепторов. Волокна тонкого и клиновидного пучков — миелинизированные,
они проводят возбуждение со скоростью 60—100 м/с. Короткие аксоны обоих пучков
устанавливают синаптические связи с мотонейронами и интернейронами своего
сегмента, длинные же направляются в продолговатый мозг. По пути они отдают
большое число ветвей к нейронам вышележащих сегментов спинного мозга, образуя,
таким образом, межсегментарные связи.
По волокнам тонкого пучка проводится возбуждение от
каудальной части тела и тазовых конечностей, по волокнам клиновидного пучка —
от краниальной части тела и грудных конечностей. В спинном мозгу оба этих пути
идут, не прерываясь и не перекрещиваясь, и оканчиваются в продолговатом мозгу у
одноименных ядер, где образуют синаптическое переключение на второй нейрон.
Отростки второго нейрона направляются к специфическим ядрам таламуса
противоположной стороны, образуя тем самым своеобразный перекрест. Здесь
они переключаются уже на третий нейрон, аксоны которого достигают нейронов IV
слоя коры больших полушарий.
Считают, что по этой системе проводится информация тонко
дифференцированной чувствительности, позволяющая определить локализацию, контур
периферического раздражения, а также его изменения во времени.
По латеральному спинно—таламическому пути проводится
болевая и температурная чувствительность, по вентральному
спинно—таламическому — тактильная. Существуют сведения, что по этим путям возможна
также передача возбуждения от проприо— и висцероцепторов. Скорость проведения
возбуждения в волокнах составляет 1—30 м/с. Спинно—таламические пути
прерываются
|
|
|
Рис. 3.19 Локализация основных восходящих путей в белом
веществе спинного мозга 1— тонкий пучок (Голля), 2— клиновидный пучок (Бурдаха), 3
— дорсолатеральный, 4 — дорсальный спинномозжечковый, 5 — вентральный
спинномозжечковый (Говерса), 6 — дорсальный спинио—таламический. |
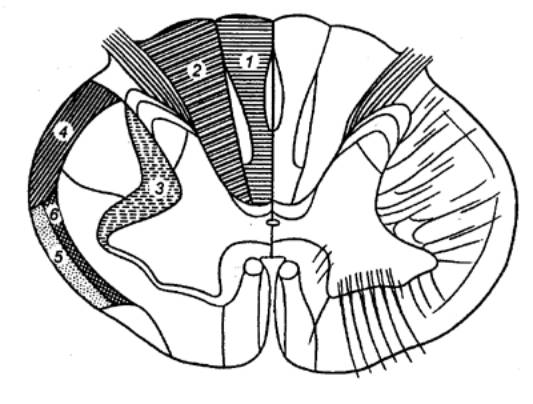
|
|
|
Рис. 3.20 Локализация основных нисходящих
путей спинного мозга 1 — латеральный корково—спинномозговой (пирамидный), 2 — красноядерно—спинномозговой (Монакова), 3 — дорсальный
преддверно—спинномозговой, 4 — оливоспинномозговой (Гельвега), 5 —
вентральный преддверно—спинномозговой, 6 — покрышечно—спинномозговой, 7 —
вентральный корково—спинномозговой (пирамидный). |
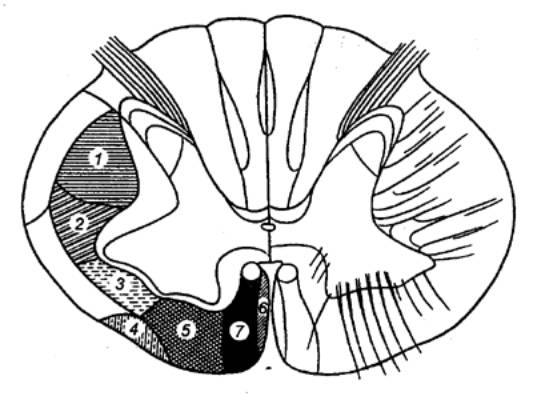
и перекрещиваются либо на уровне
сегмента, в который они только что вступили, либо вначале проходят несколько сегментов
по своей стороне, а затем переходят на противоположную. Отсюда идут волокна,
оканчивающиеся в таламусе. Там они образуют синапсы на нервных клетках, аксоны
которых направляются в кору больших полушарий.
Полагают, что по системе волокон этих путей в основном
передается информация о качественной природе раздражителей.
Дорсальный спинно—мозжечковый путь, или пучок Флексига —
филогенетически это наиболее древний чувствительный путь спинного мозга. Местом
расположения нервных клеток, аксоны которых образуют волокна этого пути,
является основание дорсального рога спинного мозга. Не перекрещиваясь, путь
достигает мозжечка, где каждое волокно занимает определенную область. Скорость
проведения по волокнам спинно—мозжечкового пути около 110 м/с. По ним проводится
информация от рецепторов мышц и связок конечностей. Наибольшего развития этот
путь достигает у копытных животных.
Вентральный спинно—мозжечковый путь, или пучок Говерса, также образуется
аксонами интернейронов противоположной стороны спинного мозга. Через
продолговатый мозг и ножки мозжечка волокна направляются к коре мозжечка, где
занимают обширные площади. Импульсы со скоростью проведения до 120 м/с идут от
сухожильных, кожных и висцерорецепторов. Они участвуют в поддержании тонуса
мышц для выполнения движений и сохранения позы.
Нисходящие проводящие пути. Эти пути (рис. 3.20) связывают
высшие отделы ЦНС с эффекторными нейронами спинного мозга. Основными из них
являются пирамидный, красноядерно—спинномозговой и ретикулярно—спинномозговой
пути.
Пирамидный путь образован аксонами клеток двигательной зоны коры больших
полушарий. Направляясь к продолговатому мозгу, эти аксоны отдают большое число
коллатералей структурам промежуточного, среднего, продолговатого мозга и
ретикулярной формации. В нижней части продолговатого мозга большая часть
волокон пирамидного пути переходит на противоположную сторону (перекрест
пирамид), образуя латеральный пирамидный путь. В спинном
мозгу он располагается в боковом
канатике. Другая часть волокон идет, не перекрещиваясь, до спинного мозга и
только на уровне сегмента, в котором оканчивается, переходит на противоположную
сторону. Это прямой вентральный пирамидный путь. Оба заканчиваются на
мотонейронах передних рогов серого вещества спинного мозга. Состав волокон
этого пути неоднороден, в нем представлены миелинизированные и
немиелинизированные волокна разного диаметра со скоростями проведения
возбуждения от 1 до 100 м/с.
Основной функцией пирамидных путей является передача импульсов для
выполнения произвольных движений. Надежность в осуществлении этой функции
повышается благодаря дублированию связи головного мозга со спинным посредством
двух путей — перекрещенного и прямого. В эволюционном ряду пирамидный тракт
развивался параллельно с развитием коры больших полушарий и достиг наибольшего
совершенства у человека.
Красноядерно—спинномозговой путь (Монакова) образован аксонами
клеток красного ядра среднего мозга. Выйдя из ядра, волокна полностью переходят
на противоположную сторону. Часть из них направляется в мозжечок и ретикулярную
формацию, другие — в спинной мозг. В спинном мозгу волокна располагаются в
боковых столбах перед перекрещенным пирамидным путем и оканчиваются на
интернейронах соответствующих сегментов. Красноядерно—спинномозговой путь несет
импульсы от мозжечка, ядра вестибулярного нерва, полосатого тела.
Основное назначение
красноядерно—спинномозгового пути — управление тонусом мышц и непроизвольной
координацией движений. В процессе эволюции этот путь возник рано. Большое
значение он имеет у животных, слабее развит у человека.
Преддверно—спинномозговой путь образован волокнами, которые
являются отростками клеток латерального пред дверного ядра (ядра Дейтерса),
лежащего в продолговатом мозгу. Этот тракт имеет наиболее древнее эволюционное
происхождение. По нему передаются импульсы от вестибулярного аппарата и
мозжечка к мотонейронам вентральных рогов спинного мозга, регулирующие тонус
мускулатуры, согласованность движений, равновесие. При нарушении целостности
этого пути наблюдаются расстройства координации движений и ориентации в
пространстве.
В спинном мозгу помимо основных
длинных имеются и короткие нисходящие пути, соединяющие между собой его
отдельные сегменты.
3.4. ПРОДОЛГОВАТЫЙ МОЗГ И МОСТ
3.4.1. Строение продолговатого мозга и моста
Продолговатый мозг и мост (варолиев
мост) представляют собой отдел головного мозга, который является
непосредственным продолжением спинного мозга. Серое вещество спинного мозга
переходит в серое вещество продолговатого мозга и сохраняет черты сегментарного
строения. Однако основная часть серого вещества распределена по всему объему
заднего мозга в виде обособленных ядер, разделенных белым веществом.
В продолговатом мозгу симметрично
заложены ядра четырех последних пар черепных нервов, осуществляющих афферентную
и эфферентную иннервацию головы и внутренних органов. В самой нижней части
ромбовидной ямки располагается двигательное ядро подъязычного нерва (XII
пара), иннервирующего мышцы языка. Двигательное ядро добавочного нерва
(XI пара) является
|
|
|
Рис. 3.21
Расположение ядер черепных нервов в стволовой части мозга 1 — добавочный нерв, 2 — подъязычный, 3 —
преддверно—улитковый, 4 — отводящий, 5 — блоковый, 6 — глазодвигательный, 7 —
тройничный, 8 — лицевой, 9 — языкоглоточный, 10 — блуждающий нерв. |
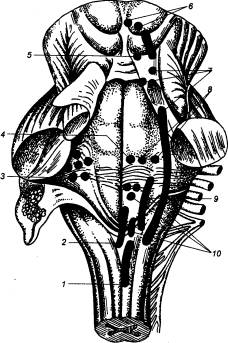
продолжением заднебокового отдела
вентрального рога спинного мозга и связано с иннервацией мышц шеи (рис. 3.21).
Довольно поверхностно лежит группа ядер
блуждающего нерва (X пара),
который является смешанным т. е. одновременно участвует в вегетативной, двигательной
и чувствительной иннервации. Соответственно функции блуждающему нерву
принадлежат три ядра. Вегетативное парасимпатическое ядро блуждающего нерва
дает парасимпатические волокна, идущие к гортани, пищеводу, сердцу, желудку,
тонкой кишке, пищеварительным железам (см. разд. 5.3). Чувствительные волокна
блуждающего нерва от многих внутренних органов в продолговатом мозгу образуют
одиночный путь (солитарныи тракт) и заканчиваются в чувствительном ядре
одиночного пути. Третье ядро блуждающего нерва — соматическое, двигательное
— лежит глубже в латеральном отделе продолговатого мозга и носит название двойного.
Нейроны этого ядра рефлекторно регулируют последовательность сокращения мышц
глотки и гортани при дыхании.
Языкоглоточный нерв (IX пара), так же как и блуждающий,
содержит двигательные, чувствительные и вегетативные волокна. Двигательные
волокна участвуют в иннервации мышц ротовой полости и глотки. Чувствительные
волокна от вкусовых почек задней трети языка, слизистой оболочки глотки входят
вместе с волокнами блуждающего нерва в одиночный путь. Отростки нейронов,
формирующих вегетативное ядро языкоглоточного нерва, идут к парасимпатическим
ганглиям, которые связаны с иннервацией секреторного аппарата слюнных желез.
На границе продолговатого мозга и моста
располагаются ядра преддверно—улиткового, или слухового, нерва
(VIII пара). Этот нерв состоит из двух частей:
улитковой и пред дверной. Волокна
улитковой части, или собственно слухового нерва, представляют собой аксоны
клеток лежащего в улитке спирального узла. Эти волокна оканчиваются в улитковых
ядрах (рис. 3.21).
Волокна преддверной части
формируются аксонами клеток узла преддверия, локализованного на дне внутреннего
слухового прохода. Эти волокна заканчиваются в трех вестибулярных ядрах моста: медиальном
(ядре Швальбе), латеральном (ядре Дейтерса) и верхнем (ядре
Бехтерева). Отростки нейронов второго порядка из этих ядер направляются к ядру
шатра мозжечка. Из латерального вестибулярного ядра, которое можно отнести к
надсегментарным образованиям, начинается преддверно—спинномозговой путь, идущий
в спинной мозг.
В латеральном отделе ретикулярной
формации моста расположено ядро лицевого нерва (VII пара). Этот нерв
является смешанным. Афферентные чувствительные волокна лицевого нерва передают
информацию от вкусовых рецепторов передней части языка. Вегетативные волокна
участвуют в иннервации подчелюстных и подъязычных слюнных желез. Эфферентные
волокна лицевого нерва имеют концевые разветвления в мимической мускулатуре
лица.
Ядро отводящего нерва (VI пара) заложено поверхностно в
задней половине моста. Моторные волокна этого нерва иннервируют мышцы,
двигающие глазное яблоко, чувствительные волокна связаны с проприоцепторами
этих мышц.
И наконец, последняя пара черепных
нервов, относящаяся к заднему мозгу, — это тройничный нерв (V пара).
Этот нерв смешанный и обладает двигательными и чувствительными ядрами. Двигательное
ядро тройничного нерва расположено в верхнем отделе покрышки моста. Нейроны
этого ядра участвуют в иннервации жевательных мышц, мышцы нёбной занавески и
мышцы, напрягающей барабанную перепонку. Длинное, простирающееся через весь
мост чувствительное ядро спинномозгового пути тройничного нерва содержит
клетки, к которым подходят афферентные аксоны от рецепторов кожи лица,
слизистой оболочки носа, зубов, надкостницы костей черепа (см. рис. 3.21).
В сером веществе продолговатого
мозга расположены ядра чувствительных путей задних столбов спинного мозга. Это тонкое
и клиновидное ядра (ядра одноименных пучков), являющихся путями, которые
проводят информацию от тактильных и проприоцепторов, сигнализирующих о
положении тела в пространстве. Кроме того, внутри заднего мозга расположено заднее
ядро трапециевидного тела (верхнее оливное ядро), включенное в восходящий
путь, который передает акустическую информацию (см. разд. 4.5.3). По
латеральной поверхности продолговатого мозга располагается продолжение боковых
столбов спинного мозга.
Центральная часть продолговатого
мозга занята ретикулярной, или сетевидной, формацией (веществом),
которая представляет собой скопление нервных клеток, напоминающее по принципу
организации нервные сети кишечнополостных. Ретикулярные нейроны распределяются
и диффузно, и группируются в ядрах, среди которых можно выделить наиболее
крупные: гигантоклеточное ретикулярное, мелкоклеточное ретикулярное,
латеральное ретикулярное ядро на уровне продолговатого мозга, каудальное,
вентрикулярное и оральное ретикулярные ядра — на уровне моста. По
цитоархитектоническим особенностям в ретикулярной формации продолговатого мозга
выделяют медиальную часть с множеством нейронов крупного размера и
латеральную, включающую в себя более мелкие элементы. По своей топографии
ретикулярная формация ствола мозга является структурой, расположенной на пути
всех входных и выходных систем мозга.
К дендритам и соме ретикулярных
нейронов подходят коллатерали от аксонов сенсорных восходящих путей,
локализованных в центральной и латеральной частях ствола мозга. С другой
стороны к нейронам ретикулярной формации подходят коллатерали нисходящих
моторных путей, идущих из переднего мозга. От нейронов гигантоклеточного и
каудального ретикулярного ядра начинается нисходящий ретикулярно—спинномозговой
путь, устанавливающий прямые связи с мотонейронами спинного мозга. Ретикулярная
формация имеет связи с ядрами черепных нервов, с мозжечком, промежуточным
мозгом и через последний — с корой больших полушарий.
Таким образом, богатство связей
ретикулярной формации свидетельствует о том, что она является надсегментарной
структурой, приспособленной к регуляции деятельности различных отделов
центральной нервной системы.
3.4.2. Рефлексы продолговатого мозга
Продолговатый мозг является жизненно
важным отделом нервной системы, где замыкаются дуги целого ряда соматических и
вегетативных рефлексов. При участии ядер продолговатого мозга осуществляются цепные
рефлексы, связанные с жеванием и проглатыванием пищи. Так, например, при
акте жевания координированная деятельность жевательных мышц, языка, щек, нёба и
дна полости рта обеспечивается функцией моторных ядер тройничного и подъязычного
нервов. Когда пищевой комок касается нёба, основания языка или задней стенки
глотки, возникает цепной рефлекс глотания. От рецепторов задней трети языка,
слизистой оболочки глотки возбуждение по чувствительным волокнам
языкоглоточного нерва поступает в продолговатый мозг к центру глотательного
рефлекса. Из этого центра запрограммированные в нервной системе моторные
команды по двигательным волокнам подъязычных, тройничных, языкоглоточных и
блуждающих нервов идут к мышцам полости рта, глотки, гортани и пищевода. В
результате координированной деятельности этих мышц совершается акт глотания с
одновременным перекрытием входов в носоглотку и гортань.
С функцией пищеварительного тракта
связаны многие вегетативные рефлексы заднего мозга. К их числу относится,
например рефлекторная регуляция секреции слюнных желез, которая осуществляется
с участием ядер лицевого и языкоглоточного нервов, содержащих парасимпатические
нейроны. Вегетативное, парасимпатическое ядро блуждающего нерва включено в
систему рефлекторной регуляции дыхания, деятельности сердца и тонуса сосудов.
Целая группа рефлексов заднего мозга
направлена на поддержание позного тонуса. В зависимости от источника
рецепторных воздействий тонические рефлексы подразделяются на шейные и
вестибулярные, или лабиринтные.
Шейные тонические рефлексы запускаются при возбуждении
проприоцепторов мышц шеи. Впервые они описаны голландским физиологом Р.
Магнусом на кошках с перерезанным на уровне шатра мозжечка стволом мозга (см.
разд. 3.5.2). В чистом виде шейные .рефлексы можно наблюдать при разрушении
вестибулярного аппарата, дающего дополнительную информацию о положении головы в
пространстве. Как показали опыты Р. Магнуса, запрокидывание головы кошки назад
в сагиттальной плоскости вызывает возрастание тонуса мышц—разгибателей передних
конечностей и одновременно ослабление экстензорного тонуса задних (рис. 3.22).
Напротив, наклон головы вниз вызывает обратный процесс, когда тонус
разгибателей передних конечностей снижается, а задних — возрастает. Нарушение
равновесия за счет поворота головы вправо или влево относительно продольной оси
вызывает компенсаторное усиление тонуса разгибателей тех конечностей, в сторону
которых повернута голова. Это так называемый рефлекс вращения по Магнусу.
Эффекторным звеном шейных рефлексов
могут быть не только мышцы конечностей, но и глазная мускулатура. Рефлекторные
влияния на глазные мышцы обеспечивают компенсаторную установку глазных яблок
при изменении положения головы. Любое отклонение головы вызывает движение
глазных яблок в противоположном направлении, и таким образом рефлекторно
сохраняется правильная зрительная ориентация животного.
Все шейные рефлексы являются
полисинаптическими. Импульсы от проприоцепторов шейных мышц идут в
соответствующие центры продолговатого мозга, а оттуда нисходящие команды
поступают или к мотонейронам спинного мозга, или к двигательному ядру
отводящего нерва и производят определенное влияние на позный тонус и на глазную
мускулатуру.
|
|
|
Рис. 3.22 Позные
рефлексы децеребрированного животного А — децеребрационная ригидность; Б, В, Г — шейные тонические
рефлексы по Магнусу; объяснение см. в тексте. |
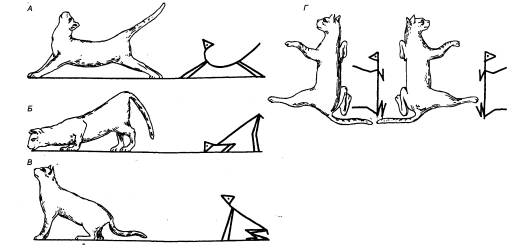
Вестибулярные рефлексы неразрывно связаны с шейными
тоническими рефлексами и в естественных условиях дополняют их. Вестибулярные
рефлексы не зависят от положения головы относительно туловища и в чистом виде
могут быть получены при фиксации головы по отношению к туловищу или при
выключении проприоцепторов шейных мышц новокаиновой блокадой.
Вестибулярные рефлексы, согласно классификации
Магнуса, подразделяются на статические и статокинетические. Статические
рефлексы связаны главным образом с возбуждением рецепторов преддверия
перепончатого лабиринта и обеспечивают поддержание позы и равновесия тела при
самых разнообразных его статических положениях в пространстве. Так, например,
если кошку с фиксированной относительно туловища головой поворачивать в
пространстве, то в зависимости от положения головы будет наблюдаться различное
распределение позного тонуса. В том случае, когда угол между плоскостью
горизонта и плоскостью ротовой щели животного составляет 45°, разгибательные
мышцы напрягаются максимально. Если же указанный угол равен 135°, наблюдается
наименьшее значение тонуса разгибателей.. Между крайними положениями имеется ряд
постепенных переходов в степени развития экстензорного тонуса. Это пример
тонического вестибулярного рефлекса положения, который осуществляется при
участии нейронов латерального вестибулярного ядра (Дейтерса) и идущего от него
в спинной мозг преддверно—спинномозгового пути.
К числу статических вестибулярных
рефлексов относят рефлексы выпрямления, направленные на переход животного из
неестественной позы в обычное для него положение. В качестве наглядного примера
можно привести переворачивание кошки в воздухе при падении спиной вниз. За
очень короткое время падения животное успевает занять нормальное по отношению к
гравитационному полю положение и упасть сразу на все четыре лапы. В процессе
переворачивания выпрямительные рефлексы совершаются в определенной последовательности.
Сначала за счет вестибулярного выпрямительного рефлекса восстанавливается
нормальное положение головы — мордой вниз. Затем изменение положения головы
возбуждает проприоцепторы шейных мышц и они запускают шейный выпрямительный
рефлекс, в результате которого вслед за головой туловище также возвращается в
нормальное положение.
Таким образом, в естественных
условиях вестибулярные рефлексы выпрямления дополняются шейными, причем ведущую
роль в их осуществлении играет положение головы, где расположены дистантные
рецепторы.
Следующая группа вестибулярных
рефлексов — статокинетические — характеризуется тем, что она направлена
на поддержание позы при изменении скорости движения животного. Эти рефлексы
связаны с возбуждением рецепторов полукружных каналов, которое имеет место при
наличии в каналах тока эндолимфы. Обычно направленность статокинетического
рефлекса зависит от того, какой полукружный канал раздражается при ускорении.
Например, при замедлении поступательного движения возбуждаются рецепторы
сагиттального канала и, соответственно, рефлекторные изменения мышечного тонуса
будут наклонять туловище вперед, как бы сохраняя равномерное движение.
Ускорение при вращении тела в
горизонтальной плоскости возбуждает рецепторы горизонтального полукружного
канала и вызывает рефлекторную реакцию глазодвигательного аппарата — горизонтальный
нистагм. Суть этой реакции состоит в том, что в момент ускорения вращения
глаза движутся в сторону, противоположную направлению вращения. Затем,
достигнув крайнего отклонения, глаза быстро перемещаются обратно в направлении
вращения и таким образом в поле зрения попадает другой участок пространства.
Нистагм способствует сохранению нормальной зрительной ориентации и обычно
используется в диагностических целях для проверки нормального функционирования
вестибулярного аппарата. Плоскость нистагма совпадает с плоскостью ускорения, в
связи с чем кроме горизонтального нистагма можно встретить вертикальный,
диагональный и круговой нистагмы (см. разд. 4.8.4).
К числу статокинетических
вестибулярных рефлексов относятся также и так называемые лифтные рефлексы,
которые проявляются в увеличении тонуса мышц разгибателей при линейном
ускорении вверх и в повышении тонуса сгибателей при линейном ускорении вниз.
3.4.3. Функции ретикулярной формации
стволовой части мозга
В ретикулярной формации
продолговатого мозга сосредоточены центры, играющие важную роль в регуляции
висцеральных функций. Это в первую очередь дыхательный центр,
локализованный в медиальной части ретикулярной формации продолговатого мозга.
Еще в прошлом столетии было установлено, что повреждение каудальной части дна
IV желудочка приводит к остановке дыхания (укол Флуранса). Позднее Н. А.
Миславский (1885) выделил в дыхательном центре две функционально различные
части: инспираторную и экспираторную (или вдыхательную и выдыхательную).
Использование методов микростимуляции и микроэлектродной регистрации активности
одиночных нейронов позволило уточнить представления о локализации частей
дыхательного центра.
Установлено, что инспираторная часть
расположена более вентрально, чем экспираторная. Активность одиночных нейронов
дыхательного центра может коррелировать с фазами дыхательного цикла. Этот
признак позволяет выделить инспираторные нейроны, которые генерируют потенциалы
действия в начальную фазу вдоха, и экспираторные нейроны, разряд которых
приурочен к фазе выдоха. Наибольшее скопление инспираторных нейронов обнаружено
около одиночного (солитарного) пути. В области двойного ядра локализованы
инспираторные и экспираторные нейроны. Определенная мозаичность — в
расположении дыхательных нейронов свидетельствует о том, что деление
дыхательного центра на две части — вдыхательную и выдыхательную — более
справедливо в функциональном смысле, чем в анатомическом (см. разд. 9.4.1).
Отличительной чертой нейронов
дыхательного центра является способность к автоматизму. Даже при
отсутствии афферентных воздействий активность этих нейронов характеризуется
периодичностью, которая определяется спецификой ионных механизмов их клеточной
мембраны. Периодичность разрядов дыхательных нейронов может быть обусловлена
также наличием взаимных тормозных связей между инспираторными и экспираторными
нейронами. Тормозные связи создают реципрокность разрядов дыхательных нейронов,
когда появление активности инспираторных нейронов сопровождается торможением
разрядов экспираторных и наоборот. В результате реципрокного взаимодействия
дыхательных нейронов происходит смена фаз дыхательного цикла.
Вместе с тем смена фаз дыхательного
цикла может осуществляться рефлекторно за счет афферентных влияний, которые
модулируют дыхательную ритмику. При интенсивном вдохе и растяжении легочной
ткани импульсы от механорецепторов легких по афферентным волокнам блуждающего
нерва приходят в дыхательный центр и вызывают рефлекторное торможение
инспираторных нейронов, одновременно возбуждая экспираторные (рефлекс
Геринга—Брейера). Импульсы от экспираторных нейронов по
ретикулярно—спинномозговому пути достигают исполнительных моторных центров
спинного мозга и стимулируют начало выдоха.
Периодичность в работе дыхательного
центра продолговатого мозга может обеспечиваться также за счет регулирующих
влияний со стороны пневмотаксического центра. Этот центр находится в
области моста и также состоит из двух типов нейронов — инспираторных и
экспираторных, которые не имеют жесткой приуроченности своих разрядов к фазам
дыхательного цикла. Пневмотаксический центр как регулятор периодически
затормаживает инспираторную часть дыхательного центра и стимулирует
экспираторные нейроны, осуществляя таким образом прекращение вдоха и начало
выдоха.
В регуляции функций дыхательного
центра могут принимать участие и более высокие этажи головного мозга. Так,
например, эмоциональные реакции человека связаны с изменением периодичности в
работе дыхательного центра, вызванным импульсацией промежуточного мозга и
лимбической коры. Установлено, что электрическое раздражение орбитальной коры
может вызвать полную остановку дыхания у человека. Регулирующие воздействия
головного мозга обеспечивают произвольную регуляцию дыхания, его коррекцию при
разнообразных изменениях жизнедеятельности.
Таким образом, регуляция
дыхательной функции осуществляется иерархической системой нервных центров,
расположенных на разных этажах нервной системы и связанных единством действия.
Другим жизненно важным центром
ретикулярной формации продолговатого мозга является сосудодвигательный
центр, локализация которого впервые была определена Ф. В. Овсянниковым
(1871). Сосудодвигательный, или вазомоторный, центр занимает обширную область
продолговатого мозга, простираясь в дорсолатеральном направлении от дна IV
желудочка до пирамид (см. разд. 5.5.2). Перерезка спинного мозга на уровне
шейных позвонков вызывает у спинальных животных резкое снижение сосудистого
тонуса и нарушение регуляторных реакций, связанных с изменением просвета вен и
артерий.
Локальная стимуляция ростральных
частей сосудодвигательного центра вызывает увеличение сосудистого тонуса,
повышение кровяного давления и тахикардию. Стимуляция каудального отдела
сосудистого центра, напротив, вызывает расширение сосудов, падение кровяного
давления и брадикардию.
Микроэлектродная регистрация
электрической активности одиночных нейронов сосудистого центра показала наличие нервных клеток, изменяющих
свою фоновую активность синхронно с флуктуациями артериального давления. Среди
них можно выделить нейроны, частота разрядов которых возрастает при повышении
артериального давления, нейроны, частота фоновой активности которых, напротив,
снижается при увеличении артериального давления, и, наконец, нейроны, частота импульсации
которых меняется в соответствии с рабочим циклом сердца. Свойства нейронов
сосудистого центра менее изучены, чем у нейронов дыхательного центра. Пока
неясно, можно ли дифференцировать сосудодвигательные нейроны на
«сосудосуживающие» и «сосудорасширяющие». Нерешенным остается также вопрос о
наличии тормозного взаимодействия между нейронами сосудодвигательного центра.
Функциональная организация
сосудодвигательного центра имеет определенную специфику в плане проекций его
нисходящих путей. Эфферентные волокна от его нейронов спускаются в грудной
отдел спинного мозга, но заканчиваются там не на мотонейронах, как в случае
нисходящих систем дыхательного центра, а на преганглионарных нейронах
симпатической нервной системы.
Таким образом, сосудистый тонус регулируется
не антагонистическими нисходящими воздействиями, а только одной симпатической
сосудосуживающей системой. Активное состояние этой системы вызывает
вазоконстрикцию, а торможение — вазодилатацию (см. разд. 5.4, 8.6). Исключение
из этого принципа составляют только некоторые сосуды, имеющие двойную
иннервацию — симпатическую и парасимпатическую (сосуды половых органов и
слюнных желез).
Рефлекторные влияния на нейроны
сосудодвигательного центра осуществляются при возбуждении хемо— и
механорецепторов, локализованных в сосудистой стенке. Афферентные волокна от
этих рецепторов в составе блуждающего и языкоглоточного нервов достигают
продолговатого мозга. Возбуждение механорецепторов дуги аорты, каротидного
синуса при повышении артериального давления вызывает торможение активности
сосудодвигательного центра и, как следствие, рефлекторное снижение сосудистого
тонуса (рефлексы Людвига—Циона, Геринга, Бейнбриджа). Напротив, при повышении
давления в системе полых вен наблюдается усиление активности сосудистого центра
и вазоконстрикторный эффект. Тонус сосудистого центра может изменяться и При
возбуждении хеморецепторов сосудистой стенки, возникающем при изменениях
химического состава крови.
Следует отметить, что деятельность
сосудодвигательного центра сочетается с функцией моторного ядра блуждающего
нерва — двойного ядра, снижающего в норме частоту сердечных сокращений. В связи
с этим при вазоконстрикторном эффекте одновременно увеличивается частота
сердечных сокращений и, наоборот, при вазодилатации наблюдается замедление
сердечной ритмики.
Для ретикулярной формации стволовой
части мозга характерны не только вегетативные регулирующие функции, но и
участие в нисходящем контроле деятельности двигательных центров спинного мозга.
В 1862 г. И. М. Сеченов установил угнетение
спинальных рефлексов при раздражении стволовой части мозга. Это было
открытием центрального торможения и одновременно открытием ретикулоспинальной
системы. Однако механизм этого тормозного процесса удалось раскрыть только в
50—е гг. нашего столетия после работ американского нейрофизиолога Г. Мегуна,
показавшего, что локальное электрическое раздражение гигантоклеточного ядра
ретикулярной формации продолговатого мозга вызывает неспецифическое торможение
сгибательных и разгибательных спинальных рефлексов. Эти неспецифические
супраспинальные влияния по ретикулярно—спинномозговому пути достигают
мотонейронов спинного мозга и увеличивают порог и скрытый период их ответов на
рефлекторные воздействия. Дальнейшие исследования показали, что нисходящие
влияния ретикулярной формации могут реализовываться не только за счет
постсинаптического торможения мотонейронов, но и за счет возникновения
длительных тормозных постсинаптических потенциалов в промежуточных нейронах, а
также за счет воздействия ретикулоспинальных волокон на терминали афферентных
волокон, входящих в спинной мозг.
Таким образом, ослабление
рефлекторной деятельности достигается за счет прямого действия на мотонейроны и
за счет определенного ослабления сенсорного входа в спинной мозг.
В ходе экспериментов с локальной
стимуляцией ретикулярной формации выяснилось наличие зон, дающих эффект
противоположной полярности, т. е. облегчающее влияние на спинномозговые
рефлексы. Так, например, электрическое раздражение латеральных зон
ретикулярной формации моста снижает порог и укорачивает скрытый период
спинальных рефлексов. При стимуляции медиальных ядер ретикулярной формации
заднего мозга в мотонейронах мышц—сгибателей кошки регистрируются
коротколатентные возбуждающие постсинаптические потенциалы (ВПСП).
Данный факт свидетельствует о
наличии (помимо диффузных неспецифических ретикулоспинальных проекций)
моносинаптических нисходящих путей специфического действия, которые участвуют в
нисходящем контроле деятельности спинного мозга.
Следовательно, ретикулярная формация
как один из двигательных центров стволовой части мозга может выступать не
только в роли регулятора возбудимости спинальных мотонейронов, но и принимать
участие в процессах, связанных с поддержанием позы и организацией целенаправленных
движений.
Благодаря работам Г. Мегуна и Дж.
Моруцци наряду с неспецифическими нисходящими влияниями ретикулярной формации
ствола были открыты ее восходящие, активирующие влияния на кору головного
мозга. Если через хронически вживленные электроды раздражать центральные
части ретикулярной формации ствола, то кошка, находящаяся в сонном состоянии,
пробуждается и у нее появляется ориентировочная реакция. Эта поведенческая
реакция пробуждения сопровождается характерными изменениями частотного спектра
электроэнцефалограммы, переходом от регулярных, высоковольтных колебаний
α—ритма к низковольтным колебаниям. (β—ритма (см. разд. 3.11.1).
Данная электроэнцефалографическая реакция получила название реакции
десинхронизации. Она имеет генерализованный характер и регистрируется от
обширных областей коры головного мозга.
В остром опыте перерезка стволовой
части мозга на уровне среднего мозга и, таким образом, разрушение восходящих
путей от ретикулярной формации ствола переводят животное в сноподобное коматозное
состояние (спящий мозг, по Бремеру) с соответствующими
изменениями характера электроэнцефалограммы.
Приведенные выше экспериментальные
факты послужили основанием для заключения, что ретикулярная формация является
структурой, отвечающей за состояние бодрствования, структурой, формирующей
восходящую активирующую ретикулярную систему, которая поддерживает на
определенном уровне возбудимость промежуточного мозга и коры больших полушарий.
Согласно современным представлениям, переход коры к активному состоянию связан
|
|
|
Рис. 3.23 Афферентные и эфферентные связи ретикулярной формации стволовой части
мозга |
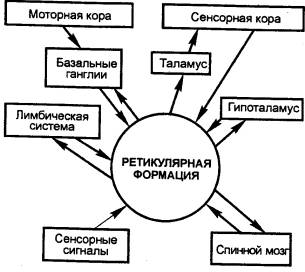
с колебаниями количества восходящих сигналов от
ретикулярной формации ствола. Количество этих сигналов зависит от поступления в
ретикулярную формацию сенсорных импульсов по коллатералям специфических
афферентных восходящих путей. Практически к ретикулярной формации приходит
информация от всех органов чувств по коллатералям от спинно—ретикулярного пути,
проприоспинальных путей, афферентных черепных нервов, от таламуса и
гипоталамуса, от моторных и сенсорных областей коры (рис. 3.23).
Микроэлектродная регистрация
электрической активности нейронов ретикулярной формации показала, что
большинство из них являются полисенсорными,
т. е. отвечают на раздражение различных модальностей (световых, звуковых,
тактильных и т. д.). Ретикулярные нейроны имеют большие рецептивные поля,
большой скрытый период и слабую воспроизводимость реакции. Эти свойства
противоположны свойствам нейронов специфических ядер и позволяют отнести
ретикулярные нейроны к категории неспецифических. Равным образом
восходящие пути ретикулярной формации в отличие от классических специфических
чувствительных проекций получили название неспецифических проекций.
Восходящие влияния ретикулярной
формации высокочувствительны к действию различных фармакологических
анестезирующих препаратов и так называемых успокаивающих средств (аминазин,
серпазил, резерпин и др.).
Использование электрофизиологических
и гистофлюоресцентных методов позволило установить медиаторную специфичность
нейронов ретикулярной формации продолговатого мозга, моста и среднего мозга.
Так, например, была изучена локализация монаминовых нейронов, которые начинают
флюоресцировать после обработки микросрезов стволовой части мозга в парах
формальдегида. Оказалось, что самая значительная группа адренергических
клеток находится в одной из областей моста, называемой «голубоватое (место)
пятно». Аксоны нейронов этой области проходят в среднем переднемозговом пучке и
имеют терминальные окончания в различных отделах головного мозга: мозжечке,
гипоталамусе, лимбической системе, коре больших полушарий.
Тела серотонинергических
нейронов расположены преимущественно в срединной области мозгового ствола — в
составе дорсального и медиального ядер шва продолговатого мозга, среднего мозга
и моста. Восходящие аксоны этих нейронов идут в гипоталамус, базальные ганглии,
лимбическую систему, кору больших полушарий.
В
настоящее время накоплен значительный фактический материал, свидетельствующий о
разнонаправленности влияний этих двух моноаминергических систем на возбудимость
различных отделов головного мозга. Серотониновая система индуцирует сон и имеет
непосредственное отношение к регуляции его медленноволновой фазы. Недостаток
серотонина в мозгу вызывает вынужденное бодрствование. Катехоламиновая система
голубоватого места, напротив стимулирует пробуждение и играет ведущую роль в
регуляции парадоксальной фазы сна. При повреждении голубоватого пятна и,
соответственно, недостатке норадреналина животные спят намного дольше, нежели
обычно.
Реципрокность моноаминергических
систем проявляется не только в процессах регуляции сна, но и в воздействии на
эмоциональную сферу организма. Например, повышение концентрации мозгового
норадреналина влечет за собой усиление стрессорного состояния животных, которое
может быть снято (купировано) введением исходного продукта синтеза серотонина —
5—окситриптофана. Равным образом разрушение ядер шва, где синтезируется
серотонин, усиливает агрессивность подопытных крыс.
Разнонаправленные влияния
моноаминергических систем мозга имеют место и в отношении регуляции
двигательной активности (серотониновая система ее снижает), и в процессах
формирования сложных форм поведения (выработка условных рефлексов). Это
свидетельствует о модулирующей функции данных систем, которые имеют обширные
связи с различными отделами головного мозга.
Другие медиаторные системы в составе
ретикулярной формации стволовой части мозга представлены холинергическими
нейронами, а также сопутствующими им глицинергическими клетками, которые чаще
всего выполняют функции тормозных интернейронов.
Следует отметить, что в составе
восходящих путей ретикулярной формации имеются активирующие и дезактивирующие
группы. По данным Дж. Моруцци, раздражение некоторых участков ретикулярной
формации заднего мозга может вызвать у животного глубокий сон и появление
α—ритма в электроэнцефалограмме. Вероятно, реципрокный принцип организации
восходящих и нисходящих проекций является общим для всей системы ретикулярной
формации.
Средний мозг расположен кпереди от
мозжечка и моста в виде толстостенной массы, пронизанной узким центральным
каналом (водопровод мозга), соединяющим полость III желудочка мозга (в промежуточном мозгу)
с IV желудочком (в продолговатом мозгу). В процессе эмбрионального развития
средний мозг формируется из среднего мозгового пузыря, боковые выпячивания
которого перемещаются латерально и образуют сетчатку глаза, которая структурно
и функционально представляет собой вынесенный на периферию нервный центр
среднего мозга.
3.5.1. Морфофункциональная организация среднего мозга
На поперечном срезе (рис. 3.24)
дорсальная поверхность среднего мозга занята пластинкой крыши, часто
называемой пластинкой четверохолмия, или четверохолмием, состоящей из двух пар
возвышений: верхних и нижних. Верхние холмики (двухолмие) играют роль
зрительного подкоркового центра и служат местом переключения зрительных путей,
идущих к латеральным коленчатым телам промежуточного мозга. У низших
позвоночных (рыб и амфибий) ростральные (верхние) холмики достигают очень
больших размеров и выполняют роль высшего зрительного центра, так как здесь
заканчивается большая часть волокон зрительного тракта.
У птиц и рептилий в среднем мозгу от
зрительных путей ответвляются немногочисленные коллатерали, идущие к
латеральным коленчатым телам промежуточного мозга. Наконец, у млекопитающих
большинство путей зрительного тракта заканчивается на нейронах коленчатых тел и
только часть из них заходит в ростральные холмики.
|
|
|
Рис.
3.24 Средний мозг А — поперечный разрез; Б—продольный разрез; 1—5 — волокна:
1 — передние корково—мостовые, 2 — корково—ядерные, 3 — латеральные
корково—спинномозговые, 4 — передние корково—спинномозговые, 5 — задние корково—мостовые,
6—водопровод среднего мозга, 7 — каудальный холмик, 8 — ретикулярная
формация, 9 — медиальная петля, 10— латеральная петля, 11— черное вещество, 12 — красное ядро.
|
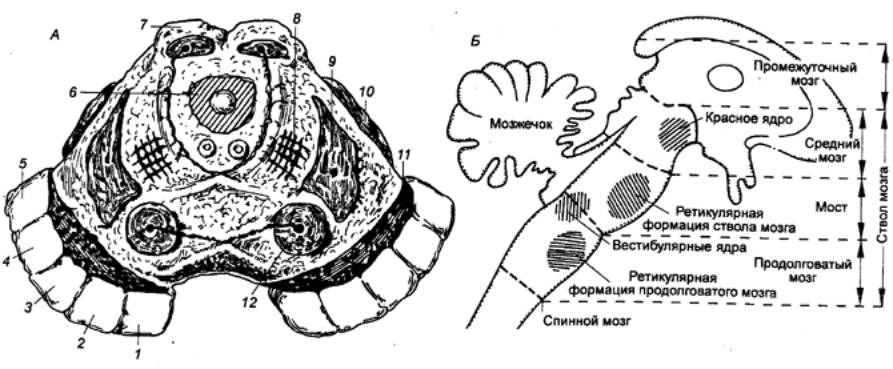
Нижние (каудальные) холмики (двухолмие) в процессе
филогенетического развития формируются у наземных животных (рептилий и птиц) в
связи с развитием органа слуха и служат местом переключения слуховых путей, а
также афферентных волокон от вестибулярных рецепторов. Каудальные (нижние)
холмики выполняют функцию подкоркового слухового центра.
Пластинка крыши и лежащие
вентральное клеточные слои вплоть до водопровода среднего мозга формируют так
называемую крышу среднего мозга, или тектальную область, которая
обладает довольно сложной цитоархитектоникой. Большие нейроны веретенообразной
формы расположены здесь слоями, общее число которых достигает 14. Ветвящиеся
дендриты и мощные аксоны этих клеток ориентированы в вертикальной плоскости по
отношению к поверхности мозга. Аксоны тектальных нейронов идут к ретикулярной
формации, к двигательным ядрам стволовой части мозга и в спинной мозг, формируя
покрышечно—спинномозговой путь. Таким образом, сама структура крыши среднего
мозга создает предпосылку для его участия в анализе сенсорной информации и в
регуляции движений.
Данные, полученные при регистрации
импульсной активности тектальных нейронов, позволяют дифференцировать их на
группы по способности реагировать на различные параметры сенсорных раздражении
(смена света и темноты, перемещение светового источника). Эфферентные
воздействия тектальных нейронов реализуются в форме ряда жизненно
важных безусловных рефлексов. К числу таких рефлексов можно отнести
сторожевой рефлекс при внезапной подаче светового или звукового раздражителей —
рефлекс, вызывающий усиление тонуса мышц сгибателей. В пластинке крыши
осуществляются замыкание ориентировочных, зрительных и слуховых рефлексов
(поворот головы к источнику раздражения, рефлекторная установка на звук
внешнего уха), оборонительных рефлексов. Все эти автоматические реакции
относятся к категории генетически запрограммированных реакций организма, важных
для сохранения вида.
В координации движений участвуют и
другие структуры среднего мозга. нейтральнее водопровода среднего мозга в виде
двух толстых валиков расположены ножки мозга, которые, расходясь кпереди,
вступают в полушария конечного мозга. На поперечном срезе ножки мозга
разделяются пигментированной прослойкой на две части: покрышку среднего
мозга (тегментум) и основание ножек мозга.
Пигментированная прослойка состоит
из нейронов, богатых пигментом меланином, и носит название черного вещества
(Земмерринга). Это филогенетически древнее образование относится к
экстрапирамидной системе регуляции двигательной активности и функционально связано
с лежащими в основании полушарий переднего мозга базальными ядрами (ганглиями)
— полосатым телом и бледным шаром (см. разд. 3.9).
В 60—е гг. нашего столетия было
установлено, что нейроны черного вещества имеют дофаминергическую природу,
т. е. способны синтезировать медиатор катехоламинового ряда — дофамин. Аксоны
этих нейронов подходят к полосатому телу, также содержащему в значительном
количестве дофамин. Дальнейшие исследования показали, что повреждение черного
вещества, вызывающее дегенерацию дофаминергических путей к полосатому телу,
связано с тяжелым неврологическим заболеванием — дрожательным параличом
(болезнью Паркинсона).
Паркинсонизм проявляется в нарушении
тонких содружественных движений, функций мимической мускулатуры и в появлении
непроизвольных мышечных сокращений, или тремора. Этот болезненный синдром может
быть снят при введении L—диоксифениланина — вещества, из которого синтезируется
дофамин в организме. Таким образом, восполняя дефицит медиатора, стало возможно
купировать неврологическое заболевание и вместе с тем привести фактические
доказательства роли черного вещества среднего мозга в сенсомоторной координации
движений.
В покрышке среднего мозга залегают
различные функционально значимые ядра. Наиболее крупным из них является парное красное
ядро (Штиллинга), представляющее собой удлиненное образование, которое
расположено между черным веществом и окружающим водопровод среднего мозга
центральным серым веществом. Красные ядра являются важным промежуточным центром
проводящих путей стволовой части мозга. В них заканчиваются волокна
экстрапирамидной системы, идущие от базальных ядер конечного мозга, а также
волокна, идущие из мозжечка.
Аксоны крупноклеточной части
красного ядра дают начало нисходящему красноядерно—спинномозговому пути
(Монакова), заканчивающемуся на мотонейронах передних рогов спинного мозга.
Этот тракт является конечным звеном древней экстрапирамидной системы,
объединяющей влияния переднего мозга, мозжечка, вестибулярных ядер и
координирующей работу двигательного аппарата.
Часть аксонов клеток, локализованных
в красном ядре, заканчивается на нейронах ретикулярной формации среднего мозга.
Она расположена несколько дорсальное красного ядра и представляет собой
продолжение ретикулярной формации заднего мозга. Наряду с активирующей
функцией, механизм которой разбирался в предыдущем разделе, ретикулярная
формация среднего мозга играет важную роль в регуляции работы
глазодвигательного аппарата.
В рефлекторной регуляции глазных
движений принимают также участие двигательные ядра глазодвигательного
(III пара) и блокового (IV пара) черепных нервов, расположенные в
покрышке под дном водопровода среднего мозга. Кпереди от ядра
глазодвигательного нерва лежит добавочное ядро глазодвигательного нерва
(ядро Даркшевича), от которого начинается медиальный продольный пучок среднего
мозга, связывающий между собой ядра глазодвигательного, блокового и
находящегося в заднем мозгу отводящего нервов, образуя из них единую
функциональную систему, регулирующую сочетанные движения глаз.
Под ядром глазодвигательного нерва
лежит непарное вегетативное парасимпатическое ядро глазодвигательного нерва
(Якубовича, или Вестфаля—Эдингера), нейроны которого посылают отростки в
периферический ресничный ганглий (см. разд. 5.1.2). Постганглионарные нейроны
ресничного ганглия иннервируют мышцы радужной оболочки, регулирующей диаметр
зрачка, и мышцы ресничного тела, изменяющие кривизну хрусталика. Рефлекторные
воздействия нейронов ресничного ганглия находятся в соответствии с
деятельностью соматических глазодвигательных ядер. Как правило, кривизна
хрусталика изменяется сопряженно с изменением угла сведения глазных осей.
Средний мозг является не только
местом замыкания многих жизненно важных рефлексов, но и выполняет существенную проводниковую
функцию. Отделенное от покрышки черным веществом основание ножек мозга
состоит исключительно из нисходящих путей, соединяющих кору больших полушарий с
мостом и спинным мозгом. В их числе находятся оба пирамидных тракта, по которым
распространяются прямые влияния коры на мотонейроны спинного мозга.
3.5.2. Участие среднего мозга в
регуляции движений и позного тонуса
Среднему мозгу принадлежит важная
роль в регуляции глазных движений. Двигательный аппарат глаза состоит из
шести наружных глазных мышц, которые иннервируются тремя черепными нервами (см.
разд. 4.8.4). Нейроны ядра глазодвигательного нерва иннервируют медиальную,
нижнюю и верхнюю прямые мышцы глаза, нижнюю косую, а также мышцу, поднимающую
верхнее веко. Блоковый нерв иннервирует верхнюю косую мышцу, а отводящий нерв —
латеральную прямую мышцу глаза. С помощью этого двигательного аппарата глаза
могут производить горизонтальные, вертикальные и вращательные движения. При
свободном рассматривании предметов, при чтении наши глаза совершают быстрые микросаккады
(микроскачки), из одной точки фиксации в другую. Микросаккады чередуются
периодами фиксации глаза, продолжающимися от 0,15 до 2 с.
Горизонтальные движения глаза зависят от содружественной
работы латеральной и медиальной прямых мышц глаза. Нейрофизиологическими
исследованиями установлено, что степень возбуждения мотонейронов,
локализованных в ядрах отводящего и глазодвигательного нервов, идущих к этим
мышцам, контролируется центрами ретикулярной формации моста. В этих центрах
обнаружены нейроны, которые характеризуются повышением частоты своей импульсной
активности перед началом каждой горизонтальной микросаккады. Другая группа
нейронов, напротив, прерывает свои импульсные разряды до и во время саккад.
Очевидно, характер разрядов нейронов
ретикулярного центра определяет его активирующие или, наоборот, тормозные
управляющие воздействия на мотонейроны отводящего и глазодвигательного нервов в
соответствии с принципом антагонистической иннервации. Аналогичным образом при вертикальных
движениях глаза также наблюдается антагонистическое взаимодействие между
двумя другими группами глазных мышц. При движении глаза по вертикали вверх
сокращаются нижняя косая и верхняя прямая мышцы (глазодвигательный нерв) и
одновременно расслабляются верхняя косая (блоковый нерв) и нижняя прямая
(глазодвигательный нерв) мышцы глаза.
В данном случае возбудимость
соответствующих моторных центров регулируется группой нейронов ретикулярной
формации среднего мозга, локализованной под ростральными холмиками пластинки
крыши. В этой области также обнаружены ретикулярные нейроны, увеличивающие
частоту своих импульсных разрядов перед началом вертикальных микросаккад.
Поражение ретикулярной формации среднего мозга при патологических процессах
парализует вертикальные движения глаз.
Таким образом, ретикулярная формация
среднего мозга играет важную роль в координации сокращений глазных мышц. Она
получает афферентные входы от ростральных холмиков пластинки крыши, мозжечка,
вестибулярных ядер, зрительных областей коры полушарий головного мозга.
Поступающие по этим входам сигналы интегрируются центрами ретикулярной формации
и служат для рефлекторного изменения работы глазодвигательного аппарата при
внезапном появлении движущихся объектов, при изменении положения головы, при
произвольных движениях глаз и т. д. По отношению к моторным центрам в ядрах
черепных нервов ретикулярная формация выступает как более высокий уровень
регуляции глазных движений, осуществляемой за счет возбуждающих и тормозных
влияний.
Одной из структур, ответственных за
надсегментарный контроль позного тонуса, является красное ядро среднего
мозга. Будучи составной частью организующей движения экстрапирамидной системы,
красное ядро получает входы от моторной коры, ядер мозжечка, черного вещества
среднего мозга и дает начало красноядерно—спинномозговому пути, который
как общий путь обеспечивает регуляцию тонуса скелетных мышц.
Локальное электрическое раздражение
крупноклеточной части красного ядра или самого красноядерно—спинномозгового
пути вызывает возбуждение α— и γ—мотонейронов мышц—сгибателей и
одновременно торможение мотонейронов разгибателей. В этом отношении влияние
красного ядра противоположно эффекту от раздражения преддверно—спинномозгового
пути, который начинается от латерального преддверного ядра (ЛПЯ).
Преддверно—спинномозговой путь оказывает возбуждающее действие на α— и
γ—мотонейроны разгибателей и тормозное — на мотонейроны сгибателей.
Существует точка зрения, что красное
ядро и ЛПЯ оказывают друг на друга тормозное воздействие, которое в норме
снижает тонус антигравитационных разгибательных мышц. О наличии такого
взаимодействия свидетельствует классический опыт Ч. Шеррингтона с перерезкой
стволовой части мозга. Если перерезку произвести на уровне переднего края
ростральных холмиков и таким образом отделить красное ядро от заднего мозга,
где локализовано ЛПЯ, то у кошки развивается децеребрационная ригидность.
Это состояние характеризуется резким
повышением тонуса мышц—разгибателей конечностей, спины и хвоста (опистотонус).
Оперированное животное, поставленное на лапы, сохраняет стоячее положение, так
как сгибания в суставах не происходит (см. рис. 3.22). Если плоскость перерезки
сдвинуть вперед и сохранить связь красного ядра с задним мозгом, то состояния
децеребрационной ригидности не возникает. Децеребрационная ригидность
устраняется при коагуляции ЛПЯ или при повторной перерезке стволовой части
мозга каудальнее зоны расположения вестибулярных ядер.
Все эти факты указывают на то, что
основной причиной децеребрационной ригидности является снятие при перерезке
тормозных влияний со стороны красного ядра на ЛПЯ. При этом начинает
преобладать действие ЛПЯ, которое, как известно, возбуждает мотонейроны
мышц—разгибателей.
По представлениям Ч. Шеррингтона,
красное Ядро является не только источником, но и посредником в регуляции
разгибательных рефлексов, передавая тормозные влияния со стороны мозжечка и
моторных областей коры. Удаление мозжечка или моторной коры сопровождается
усилением тонуса разгибательных мышц.
Децеребрационная ригидность—
рефлекторное состояние, оно поддерживается сенсорными сигналами от
проприоцепторов мышц. Если перерезать дорсальные (чувствительные) корешки
спинного мозга, то спазматическое сокращение разгибательных мышц исчезает.
Очевидно, в естественных условиях красное ядро регулирует протекание
проприоцептивных рефлексов спинного мозга, подчиняя их рефлексам более высокого
порядка. Одним из путей этой регуляции является изменение возбудимости
γ—мотонейронов, контролирующих чувствительность проприоцепторов мышц.
Другой путь состоит в непосредственном воздействии на α—мотонейроны мышц—разгибателей.
Мозжечок как надсегментарная
структура появляется на ранних этапах филогенеза позвоночных, причем степень
его развития у различных животных определяется экологией и сложностью
локомоции.
3.6.1. Структурная организация и связи мозжечка
У млекопитающих мозжечок — крупный
вырост моста, состоящий из двух полушарий и непарного отдела — червя.
Со стволовой частью мозга мозжечок соединяется тремя парами ножек. Самые
толстые средние ножки как бы охватывают продолговатый мозг и,
расширяясь, переходят в мост. Ростральные ножки начинаются в зубчатых
ядрах мозжечка (см. ниже) и направляются к пластинке крыши среднего мозга.
Третья пара ножек (каудальная) спускается вниз, сливаясь с продолговатым
мозгом. Афферентные волокна, приходящие в мозжечок, преимущественно входят в
состав средних и каудальных ножек, тогда как эфферентные собраны главным
образом в ростральных ножках мозжечка.
Вся поверхность мозжечка разделяется
глубокими бороздами на доли. В свою очередь, каждая доля параллельными
бороздками разделяется на извилины;группы извилин формируют дольки
мозжечка. Каждую дольку обозначают как классическим названием (язычок,
центральная, вершина и т. д.), так и латинской нумерацией (I—Х) в соответствии с принятой
номенклатурой (рис. 3.25).
Согласно О. Ларселу, всю поверхность
мозжечка можно разделить на отделы в зависимости от характера поступающих
афферентных путей и филогенетического возраста структурных образований.
Наиболее изолированная клочково—узелковая доля (X) составляет древнюю
часть мозжечка (палеоцеребеллум), гомологичный мозжечку круглоротых. Здесь
заканчиваются проекции от вестибулярных ядер продолговатого мозга. Следующий
отдел — старая часть мозжечка, или археоцеребеллум, — включает в себя
участки червя, соответствующие ростральной доле, а также пирамиду, язычок и
околоклочок. В палеоцеребеллуме находятся проекции восходящих
спинно—мозжечковых путей, несущих информацию от мышечных рецепторов. И наконец,
третий отдел — новая часть мозжечка, или неоцеребеллум, — состоит из
появляющихся у млекопитающих полушарий и участков червя, которые расположены
каудальнее первой щели. К неоцеребеллуму по путям, переключающимся в ядрах
моста, поступает афферентная импульсация от обширных областей коры больших полушарий
(лобных, теменных, височных и затылочных долей).
Полушария и червь мозжечка состоят из
лежащего на периферии серого вещества — коры. — и расположенного глубже
белого вещества — мозгового тела, в котором заложены скопления нервных
клеток, образующие ядра мозжечка. Кора мозжечка представлена тремя слоями,
каждый из которых имеет определенный набор клеточных элементов. Самый
поверхностный слой — молекулярный — состоит из параллельных волокон и
разветвлений дендритов и аксонов нейронов нижележащих слоев. В нижней части
молекулярного слоя расположены тела корзинчатых нейронов, аксоны которых
оплетают тела и начальные сегменты аксонов клеток Пуркинье (грушевидных
нейронов) (рис. 3.25). Здесь же в молекулярном слое имеется некоторое
количество звездчатых клеток.
Вентральное молекулярного слоя
находится слой грушевидных нейронов, в котором сосредоточены тела клеток
Пуркинье (рис. 3.25). Эти
крупные клетки ориентированы вертикально по отношению к поверхности коры
мозжечка. .Их дендриты поднимаются вверх и широко ветвятся в молекулярном слое.
Дендриты клеток Пуркинье содержат множество шипиков, на которых образуют
синапсы параллельные волокна молекулярного слоя. Аксоны клеток Пуркинье
спускаются к ядрам мозжечка. Часть из них заканчивается на вестибулярных ядрах.
Практически аксоны клеток Пуркинье представляют собой единственный выход из
коры мозжечка.
Под ганглиозным слоем лежит зернистый
слой, который содержит большое число тел клеток—зерен, или
гранулярных клеток. По некоторым подсчетам их число может достигать 10 млрд.
Аксоны клеток—зерен поднимаются вертикально вверх, в молекулярный слой и там
Т—образно ветвятся. Ветви идут параллельно поверхности коры и образуют синапсы
на дендритах других клеток. Здесь же в гранулярном слое лежат клетки
Гольджи, аксоны которых подходят к клеткам—зернам.
Афферентный вход к нейронному аппарату коры
осуществляется по трем системам нервных волокон. Это, во—первых, лазающие,
или ползучие, волокна, идущие из нижних олив продолговатого мозга. Нижняя
олива получает афференты от нескольких восходящих путей спинного мозга и из
центров головного мозга. Лазающие волокна широко ветвятся и подобно лианам
оплетают дендриты клеток Пуркинье, формируя на них синапсы. Вторая система
афферентных
|
|
|
Рис. 3.25 Морфофункциональная организация мозжечка А — отделы и доли мозжечка; Б — расположение связей в коре
мозжечка: I—X — доли мозжечка по номенклатуре Ларсела; 1 — ростральная доля, 2 — каудальная доля, 3 —
околоклочок, 4 —
клочково—узелковая доля, 5 — корзинчатая клетка, 6 — клетка Пуркинье, 7 — лазающее волокно, 8 — клетка—зерно, 9 —клетка внутримозжечкового ядра, 10 — клетка Гольджи, 11 — мшистое волокно; на рис. А черным цветом обозначены
структуры древней части мозжечка, серым — старой части, светлым — новой части
мозжечка. |
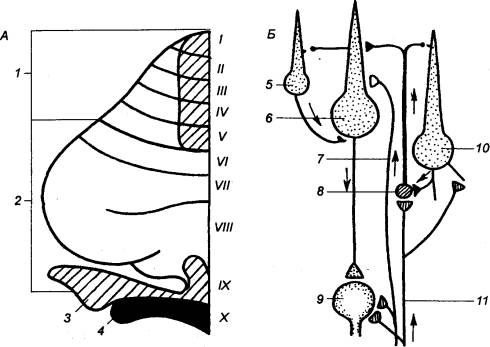
волокон — это моховидные, или
мшистые, волокна, идущие от ядер моста и оканчивающиеся на
клетках—зернах. Мшистые волокна многократно ветвятся и образуют синапсы на множестве
клеток коры мозжечка. И наконец, третья система афферентных волокон — это также
широко ветвящиеся адренергические волокна, поступающие в кору мозжечка
из голубоватого пятна в среднем мозгу. Оно представляет собой скопление из
нескольких нейронов, аксоны которых способны диффузно выбрасывать норадреналин
в межклеточное пространство. Вероятно, эти нейроны выполняют нейромодуляторную
функцию и могут изменять возбудимость нейронов, локализованных в коре мозжечка.
Нейрофизиологические исследования
Дж. Эклса показали, что корзинчатые и звездчатые клетки, которые заканчиваются
синапсами на клетках Пуркинье, вызывают в них тормозные постсинаптические
потенциалы (ТПСП) и подавление импульсной активности. Клетки Гольджи тормозят
клетки—зерна по принципу обратной связи (рис. 3.25).
Таким образом, большинство связей,
опосредованных интернейронами коры мозжечка, являются тормозными. Исключение
составляют только клетки—зерна, которые возбуждаются от моховидных волокон и
сами через Т—образно ветвящиеся аксоны активируют все остальные интернейроны
коры мозжечка. Однако конечный эффект этой активации опять—таки сводится к
торможению.
Клетки Пуркинье, которые
представляют собой выход функциональной системы, могут возбуждаться прямо через
лазащие волокна и опосредованно через моховидные волокна и клетки—зерна.
Возникающие под действием этого возбуждения разряды клеток Пуркинье, согласно
электрофизиологическим данным, вызывают в конечном счете торможение нейронов
ядер мозжечка. Эти факты свидетельствуют о том, что деятельность всей
нейрональной системы коры мозжечка сводится к торможению ядер, над которыми
кора надстроена. Очевидно, механизм этого торможения можно представить
следующим образом.
В покое клетки Пуркинье обладают
фоновой электрической активностью, которая вызывает тоническое торможение
нейронов в ядрах мозжечка. Возбуждение клеток Пуркинье через систему лазащих
или моховидных волокон приводит к увеличению частоты импульсных разрядов этих
нейронов и, как следствие, к усилению торможения ядер мозжечка. Напротив,
торможение клеток Пуркинье, вызванное звездчатыми или корзинчатыми клетками,
сопровождается растормаживанием нейронов в ядрах мозжечка. Сами же ядра
мозжечка, обладающие постоянной тонической активностью, через нисходящие пути
регулируют уровень возбудимости центров спинного мозга и мышечный тонус.
Согласно гипотезе, высказанной Дж.
Эклсом, большое количество тормозных нейронов в коре мозжечка предотвращает
длительную циркуляцию возбуждения по нейронным цепям. Любой возбуждающий
импульс, приходя в кору мозжечка, превращается в торможение примерно за 100 мс.
Так происходит как бы автоматическое стирание предшествующей информации,
которое позволяет коре мозжечка участвовать в регуляции быстрых движений.
В белом веществе мозжечка
сконцентрированы три пары ядер. В белом веществе червя близко к срединной
плоскости находится ядро шатра (Кёлликера). Нейроны этого ядра посылают
свои отростки к ЛПЯ и к ретикулярной формации продолговатого мозга и моста, где
берет свое начало ретикулярно—спинномозговой путь. Латеральное ядра шатра
находятся вставочные, или промежуточные, ядра, которые у человека
разделяется на латеральное (пробковидное) и медиальное
(шаровидное) вставочные ядра мозжечка.
От вставочного ядра аксоны идут в средний
мозг к красному ядру. Менее развитый афферентный путь от вставочного ядра идет
в промежуточный мозг к вентролатеральному ядру таламуса (см. разд. 3.7.2) — и
оттуда к двигательной коре. Латеральное всех ядер лежит наиболее крупное латеральное
(зубчатое) ядро мозжечка, от которого мощные пучки волокон направляются
к вентролатеральному ядру таламуса, и далее аксоны нейронов второго порядка
проецируются в моторные зоны коры.
К нейронам мозжечковых ядер подходят
аксоны клеток Пуркинье. Установлено, что клетки Пуркинье червя образуют прямые
связи с ЛПЯ. Это позволяет иногда относить ЛПЯ к внутримозжечковым ядрам по
функциональному принципу.
Существует определенная топография
связей коры мозжечка с его ядрами. Согласно классификации Бродала, кору
мозжечка млекопитающих можно разделить на три продольные зоны: медиальную
червячную зону, от которой аксоны клеток Пуркинье проецируются на ядро
шатра, промежуточную зону коры, связанную со вставочным ядром, и латеральную
зону коры полушарий, дающую проекции к латеральному ядру. Эта
классификация, в основу которой положены эфферентные связи мозжечка,
свидетельствует о том, что латеральные отделы мозжечка через зубчатое ядро
связаны с более высокими уровнями головного мозга.
В целом мозжечок имеет обширные
эфферентные связи со всеми двигательными системами стволовой части мозга:
кортикоспинальной, руброспинальной, ретикулоспинальной и вестибулоспинальной.
Не менее разнообразными являются и афферентные входы мозжечка.
Афферентная информация в мозжечок от
спинного мозга приходит по спинно—мозжечковым путям (дорсальному и
вентральному), ростральному спинно—мозжечковому и клиновидно—мозжечковому
путям, по спинно—оливо—мозжечковым путям. Кора больших полушарий также посылает
афферентные пути в мозжечок, среди которых наиболее важными являются
корково—ретикулярно—мозжечковый и корково—мосто—мозжечковый пути.
Мозжечок как надсегментарный орган,
входящий в систему регуляции движений, выполняет следующие важные функции:
1) регуляция позы и мышечного тонуса;
2) сенсомоторная координация позных
и целенаправленных движений;
3) координация быстрых
целенаправленных движений, осуществляемых по команде из больших полушарий. Эти
функции мозжечка наиболее удобно разбирать в соответствии с топической классификацей
его отделов, основанной на характере эфферентных связей.
Медиальная червячная зона мозжечка в
наибольшей степени связана с реализацией первой функции, т. е. с управлением тонусом,
позой и равновесием тела. В эту зону коры мозжечка, а также в клочково—узелковую
долю палеоцеребеллума поступает афферентная информация, сигнализирующая о позе
и состоянии локомоторного аппарата. После обработки этой информации из коры
мозжечка через ядро шатра корригирующие команды направляются к ЛПЯ, к
ретикулярной формации ствола и оттуда к спинальным центрам по ретикулярно— и
преддверно—спинномозговым путям.
Таким образом, вся система работает
по принципу обратной связи и обеспечивает срочную регуляцию антигравитационного
тонуса.
Удаление червячной зоны мозжечка вызывает
растормаживание ЛПЯ и как результат — усиление тонуса разгибателей животного в
состоянии децеребрационной ригидности. Электрическое раздражение червячной
зоны, напротив, приводит к ослаблению тонуса мышц разгибателей за счет
активации тормозных влияний мозжечка на ЛПЯ.
Промежуточная зона коры мозжечка,
связанная со вставочным ядром (или шаровидным и пробковидным), так же как и
предыдущая зона, имеет афферентные входы от спинномозговых путей. Однако кроме
того, через ядра моста промежуточная зона коры мозжечка получает информацию от
двигательной области коры больших полушарий. Эта информация поступает через
коллатерали корково—спинномозгового пути и сигнализирует о готовящемся
целенаправленном движении. Сопоставление приходящей по этим двум путям информации
позволяет промежуточной зоне мозжечка участвовать в координации
целенаправленных движений с рефлексами поддержания позы, в выборе позы
оптимальной для выполнения движения. Нисходящие команды от промежуточной зоны
коры мозжечка через вставочное ядро идут к красному ядру и далее по
красноядерно—спинномозговому пути к моторным центрам спинного мозга.
Как уже отмечалось, латеральная,
филогенетически наиболее молодая, кора полушарий мозжечка дает эфферентные
проекции к зубчатому ядру. Эта латеральная зона через корково—мосто—мозжечковый
путь получает афферентный вход от различных ассоциативных зон коры больших
полушарий. По этим афферентным путям в кору полушарий мозжечка поступает
информация о замысле движения. В полушариях и зубчатом ядре мозжечка эта информация
преобразуется в программу движения, которая по мозжечково—таламо—корковому пути
поступает в двигательные зоны коры больших полушарий. В дальнейшем двигательный
акт реализуется за счет нисходящих команд, идущих от двигательной коры в
спинной мозг по корково—спинномозговому пути. Кроме того, от зубчатого ядра
мозжечка через красное ядро могут распространяться прямые нисходящие
воздействия к спинальным центрам.
Эта сложная система регуляции
движений с наличием обратных связей между мозжечком и корой больших полушарий
позволяет полушариям мозжечка участвовать в организации быстрых
целенаправленных движений, протекающих без учета информации, идущей от
восходящих сенсорных путей спинного мозга. Такие движения могут встретиться в
спортивной практике, при игре на музыкальных инструментах и при некоторых
других видах деятельности.
Экспериментальные исследования
функций мозжечка показали, что этот отдел головного мозга участвует не только в
регуляции движений, но и включен в систему контроля висцеральных функций.
Л. А. Орбели было установлено, что раздражение мозжечка вызывает целый ряд
вегетативных рефлексов, таких, например, как расширение зрачков, повышение
артериального давления и т. д. Удалению мозжечка сопутствуют нарушения
сердечнососудистой деятельности, дыхания, моторной и секреторной функций
желудочно—кишечного тракта. Электрофизиологическими методами в мозжечке
обнаружены висцеральные проекции внутренних органов. Так, например, при
раздражении интероцепторов в коре мозжечка регистрируются вызванные потенциалы.
Все эти экспериментальные факты
позволили Л. А. Орбели считать мозжечок млекопитающих не только регулятором
моторной деятельности, но и адаптационно—трофическим органом, который
определяет возбудимость вегетативных и соматических центров. Однако до сих пор
остается неясным вопрос о том, какое место занимает мозжечок в иерархии
структур, регулирующих висцеральные функции. По всей вероятности, модулирующие
влияния мозжечка могут быть опосредованы связями с ретикулярной формацией
ствола и гипоталамусом, который является главным подкорковым центром регуляции
висцеральных функций.
Основные функции мозжечка определяют
и характер патологических симптомов при нарушении его деятельности.
Согласно классическим представлениям Л. Лючиани, сформулированным еще в конце
XIX века, при частичном и общем поражении мозжечка наблюдаются симптомы:
атония, астения и астазия.
Атония характеризуется ослаблением
мышечного тонуса. У собак и кошек после удаления мозжечка наблюдается начальное
повышение тонуса мышц—разгибателей, причем животные находятся в позе, сходной с
состоянием децеребрационной ригидности. Однако через несколько суток после
операции у животных гипертонус разгибателей сменяется гипотонией. Аналогичная
операция у приматов сразу вызывает падение мышечного тонуса и как следствие
неспособность поддерживать определенную позу. Обычно атония сопровождается астенией,
которая характеризуется слабостью и быстрой утомляемостью мышц. Очевидно,
атония и астения связаны в первую очередь с выпадением функции мозжечка контролировать
познотонические рефлексы.
Третий симптом, описанный Л.
Лючиани, — астазия — проявляется в способности мышц производить
колебательные и дрожательные движения. Мышечный тремор особенно выражен в
начале и конце движения, что в значительной мере препятствует завершению
целенаправленного движения. Этот симптом вызывается нарушением сенсомоторной
координации позных и целенаправленных движений.
Более поздние исследования,
проведенные на млекопитающих, показали, что удаление мозжечка пагубно
сказывается на выполнении произвольных движений. При этом более всего страдают
содружественные движения — появляются симптомы асинергии. В результате
асинергии происходит как бы распад программы движения и целостное движение
теперь состоит не из одновременных содружественных актов, а из
последовательности ряда простых движений. Так, например, касание рукой кончика
носа мозжечковый больной осуществляет в три приема. Асинергия сочетается с дисметрией,
или утратой соразмерности движения. Движения становятся преувеличенно размашистыми
и теряют свою точность, в результате чего цель достигается только после
многократных ошибок. У мозжечковых больных деформируется походка— атаксия.
Атаксическая походка характеризуется широко расставленными ногами и избыточными
движениями, из—за которых больного как пьяного «бросает» из стороны в сторону.
Нарушение координации быстрых
целенаправленных движений при удалении или травмах мозжечка может вызвать адиадохокинез
— неспособность выполнять быструю последовательность противоположных
движений, например сгибание и разгибание пальцев.
Следует отметить, что у
млекопитающих, перенесших травму мозжечка, со временем наступает довольно
эффективная компенсация его функций. По всей вероятности, эта компенсация
осуществляется за счет коры больших полушарий, которая обладает двусторонними
связями с мозжечком. Этот факт свидетельствует о высокой пластичности мозговых
систем регуляции движений.
3.7.1. Структура промежуточного
мозга
Промежуточный мозг в процессе эмбриогенеза развивается
из переднего мозгового пузыря и образует стенки III желудочка. Топографически и
функционально промежуточный мозг подразделяется на эпиталамус, таламус и
гипоталамус.
Эпиталамус (надталамическая область), или
надбугорье, состоит из поводка и железы внутренней секреции — шишковидного тела
(эпифиза), которые формируют верхнюю стенку III желудочка. Таламус (От
греч. thalamos — покой, опочивальня), или зрительный бугор,
представляет собой состоящее из скопления серого вещества объемистое тело
яйцевидной формы. Нижней и латеральной поверхностью таламус сращен с соседними
частями мозга. Медиальная поверхность зрительного бугра образует боковую стенку
полости III желудочка. Таламус является крупным подкорковым образованием, через
которое в кору больших полушарий проходят разнообразные афферентные пути.
Дно III желудочка формирует группа
структур, которые объединяют под названием гипоталамуса (подбугорья).
Гипоталамус содержит большое количество ядер и является центром регуляции
висцеральных функций организма (см. разд. 3.7.3).
3.7.2. Морфофункциональная
организация таламуса
Нервные клетки таламуса группируются
в большое количество ядер (до 40), которые топографически разделяют на
переднюю, заднюю, срединную, медиальную и латеральную группы. По функции
таламические ядра можно дифференцировать на специфические, неспецифические,
ассоциативные и моторные.
В специфических, или проекционных,
ядрах таламуса происходит синаптическое переключение сенсорной информации с
аксонов восходящих афферентных путей на следующие, конечные нейроны, отростки
которых идут в соответствующие сенсорные проекционные области коры больших
полушарий. Повреждение специфических ядер приводит к необратимому выпадению
определенных видов чувствительности. Эти экспериментальные факты
свидетельствуют о том, что специфические ядра являются передаточной станцией на
пути афферентных импульсов от периферических рецепторов к коре больших
полушарий.
Среди основных проекционных ядер
таламуса можно выделить заднее вентральное ядро, которое является
специфическим ядром соматосенсорной системы. Оно разделяется на две части — заднелатеральное
вентральное ядро, к которому подходят восходящие волокна
спинно—таламического тракта и системы медиальной петли, несущие информацию от
кожных рецепторов туловища, проприоцепторов мышц и суставного аппарата, и заднемедиальное
вентральное ядро, к которому подходят соответствующие пути от ядер
тройничного нерва, осуществляющего иннервацию лицевой части головы.
Микроэлектродные исследования
нейронов заднего вентрального (вентро—базального) комплекса показали, что
данное ядро, как и прочие специфические ядра, организовано по топическому
принципу. Суть этого принципа состоит в том, что каждый нейрон активируется
раздражением рецепторов определенного участка кожи, причем смежные участки
туловища проецируются на смежные части вентробазального комплекса.
Специфичность данных нейронов
проявляется также в том, что каждый из них возбуждается одним типом рецепторов.
Соматотопический принцип организации сохраняется и на более высоком Уровне в
соматосенсорной проекционной области коры больших полушарий (постцентральная
извилина), с которой задний вентральный комплекс связан восходящими и
нисходящими путями.
Специфическим ядром зрительной сенсорной
системы является латеральное коленчатое тело (ЛКТ), составляющее вместе
с медиальным коленчатым телом метаталамус и имеющее прямые связи с
затылочными (зрительными) проекционными областями коры больших полушарий.
ЛКТ имеет слоистую структуру и организовано
также по топическому принципу. Аксоны, идущие в ЛКТ из зрительного тракта,
'распределяются в нем с поразительной четкостью: три слоя ЛКТ связаны с
ипсилатеральным (расположенном на той же стороне) глазом, а три остальные — с
контрлатеральным. В каждом из слоев ЛКТ аксоны зрительного тракта вступают в
синаптический контакт с четко ограниченными группами клеток. Нейроны ЛКТ
обладают концентрическими рецептивными полями, центр и периферия которых
антагонистичны и по—разному реагируют на изменение освещенности. Рецептивное
поле нейрона ЛКТ представляет собой концентрически организованную совокупность
рецепторов сетчатки, имеющую либо возбуждающий (при включении света) центр и
периферическую тормозную часть, либо, напротив, тормозный центр и возбуждающую данный
нейрон ЛКТ периферическую область.
Такая организация рецепторного поля
позволяет нейронам ЛКТ хорошо реагировать на контраст при определении границы
между темным и светлым и на суммарную яркость светового стимула.
У млекопитающих некоторые нейроны ЛКТ
обладают цветоспецифичными рецепторными полями и могут возбуждаться или,
наоборот, тормозиться в зависимости от длины волны светового стимула. Таким
образом, нейроны ЛКТ, так же как и нейроны сетчатки, принимают участие в
анализе зрительной информации. Об этом же свидетельствует наличие тормозных
процессов в нейронах ЛКТ, субстратом которых являются цепи отрицательной
обратной связи, образованные возвратными коллатералями аксонов этих клеток и
промежуточными нейронами, имеющими тормозные окончания. По всей вероятности,
возникающие в нейронах ЛКТ тормозные постсинаптические потенциалы служат для
подавления следовых эффектов и всех побочных воздействий, которые мешают
передаче сигнала через данный канал.
Восходящие пути слуховой системы,
идущие из каудальных холмиков и по волокнам латеральной петли, проецируются в
специфическое таламическое ядро — медиальное коленчатое тело (МКТ), от
которого начинается путь, достигающий первичной слуховой коры в верхней части
височных долей.
Медиальное коленчатое тело состоит
из мелкоклеточной и крупноклеточной частей и обладает тонотопической
специализацией своих нейронов. Так, например, нервные клетки мелкоклеточной
части МКТ имеют довольно узкую настройку на восприятие звуков различной высоты
и принимают участие в анализе и передаче акустической информации.
Таким образом, таламус является
посредником, в котором сходятся все раздражения от внешнего мира (кроме
запахов) и, видоизменяясь здесь, направляются к подкорковым и корковым центрам.
В специфические ядра таламуса проецируются
афференты не только от экстероцепторов и рецепторов двигательного аппарата.
Электрофизиологические исследования показали, что в заднем вентральном
комплексе таламуса имеются области проекций блуждающего и чревного нервов,
чувствительные волокна которых несут информацию от интероцепторов. В то же
время таламус, как надсегментарный центр рефлекторной деятельности, имеет связи
|
|
|
Рис.
3.26 Проекции специфических (а) и неспецифических (б) ядер таламуса в кору
больших полушарий 1 — сенсорные афферентные пути, 2 —
ретикулярно—таламические пути, 3 — проекционная зона коры, 4 — ассоциативная
зона коры. |
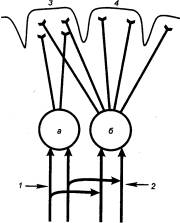
с гипоталамусом, где сосредоточены
главные вегетативные центры. Эти связи характерны для передних ядер таламуса и
создают материальную предпосылку для участия этой структуры в системе регуляции
висцеральных функций организма.
Следующую функциональную группу ядер
таламуса составляют так называемые ассоциативные ядра. В отличие от
специфических ядер они не могут быть отнесены к какой—либо одной сенсорной
системе и получают афферентные импульсы от специфических проекционных ядер. Три
ядра этой группы имеют связи с главными ассоциативными областями коры: ядро
подушки связано с ассоциативной зоной теменной и височной коры, заднее
латеральное ядро — с теменной корой, дорсальное медиальное ядро — с
лобной долей. Четвертое ядро — переднее — имеет связи с лимбической
корой больших полушарий. По—видимому, ассоциативные ядра участвуют в высших
интегративных процессах, однако их функция изучена еще недостаточно.
К моторным ядрам таламуса относится вентролатеральное
ядро, которое имеет вход от мозжечка и базальных ганглиев и одновременно
дает
проекции в моторную зону коры больших
полушарий. Это ядро включено в систему регуляции движений, и, как показал
материал клиники, разрушение некоторых его участков ослабляет симптомокомплекс
болезни Паркинсона.
Наконец, последнюю большую группу
ядер таламуса образуют неспецифические ядра, которые функционально
связаны с ретикулярной формацией ствола. К числу этих ядер относится медианная
и внутрипластинчатая группы ядер таламуса, которые получают афферентные
входы от волокон, восходящих из ретикулярной формации и, кроме того, имеет двусторонние
связи со специфическими ядрами таламуса. В отличие от специфических ядер с
локальными проекциями в коре филогенетически более древние неспецифические ядра
обнаруживают диффузные проекции во все области коры (рис. 3.26). Этой
структурной особенностью обусловлены их название и функция, которая состоит в
регуляции возбудимости и электрической активности корковых нейронов.
Доказательства влияния
неспецифических ядер таламуса на кору впервые были получены американскими
исследователями Э. Демпси и Р. Моррисоном в 1942 г. Они показали, что при
электрическом раздражении неспецифических ядер с ритмом 6—12 имп/с почти на
всей поверхности коры ипсилатерального полушария регистрируются постепенно
увеличивающиеся по амплитуде негативные волны, очень сходные с α—ритмом
электроэнцефалограммы. Эти негативные волны появляются с латентным периодом 25
мс и более на 2—3—й стимул, достигают максимальной амплитуды на 5—6—й стимул, а
затем начинают постепенно уменьшаться вплоть до полного исчезновения. Если
раздражение продолжается, то негативные волны появляются вновь в такой же
последовательности. Эта электрофизиологическая реакция была названа рекрутирующим
ответом, или реакцией вовлечения.
В отличие от первичных ответов,
которые регистрируют при раздражении специфических ядер, реакция вовлечения
характеризуется большим латентным периодом, нарастанием и снижением амплитуды и
отсутствием локальности или диффузностью вне связи с какой—либо специфической
областью коры.
Дальнейший анализ этого
электрофизиологического феномена показал, что периодическое увеличение и
уменьшение негативных волн реакции вовлечения обусловлено лучшей или худшей
синхронизацией активности таламических и корковых нейронов, а также суммацией
постсинаптических потенциалов в большем или меньшем количестве нейронов.
Уровень синхронизации может определяться динамикой возбуждающих и тормозных
процессов при таламо—кортикальных циклических взаимодействиях.
Наличие этих взаимодействий
используют для объяснения природы ритмической активности коры больших полушарий
и, в частности, происхождения α—ритма электроэнцефалограммы (см. разд.
3.11.1).
При сопоставлении функций
специфических и неспецифических ядер таламуса возникает резонный вопрос о
взаимодействии этих двух систем, которые могут влиять на одни и те же нейроны
коры больших полушарий. Как показали электрофизиологические исследования,
восходящие влияния неспецифических ядер таламуса проявляются не в вызове
разряда коркового нейрона, а в изменении его возбудимости. Неспецифические
влияния из таламуса, повышая возбудимость корковых нейронов, облегчают их
деятельность, при этом ответы корковых нейронов на импульсы, приходящие из
специфических проекционных ядер, усиливаются. Вместе с тем неспецифические
влияния могут иметь и противоположный знак и обнаруживать угнетающее действие
на разряды корковых нейронов.
Существует точка зрения, что
неспецифические ядра включены в восходящую активирующую систему и являются
посредниками между корой и ретикулярной формацией ствола, которая получает
информацию от всех органов чувств. Таким образом, неспецифические ядра передают
активирующие влияния ретикулярной формации и участвуют в поддержании
оптимального тонуса коры. Однако эта точка зрения не является общепризнанной, и
некоторые исследователи рассматривают ретикулярную формацию и неспецифические
ядра таламуса как две раздельные системы, контролирующие возбудимость корковых
нейронов.
Было бы упрощением рассматривать
таламокортикальные взаимодействия, как односторонние. Кора, в свою очередь,
может оказывать тормозные и облегчающие влияния на таламические ядра. Эти
влияния могут распространяться на специфические ядра по прямым
кортикоталамическим путям и на неспецифические через
кортикоретикулоталамические связи. Так, например, раздражение сенсомоторной
зоны коры угнетает на длительное время возникновение реакции вовлечения. Эти
факты свидетельствуют о наличии между таламусом и корой больших полушарий
двусторонних циклических связей, которые играют важную роль в интегративной
деятельности мозга. Не случайно таламокортикальная система связана с регуляцией
таких физиологически важных состояний, как смена сна и бодрствования,
сохранение сознания, развитие процессов внутреннего торможения.
Гипоталамус — филогенетически старый отдел
промежуточного мозга, который играет важную роль в поддержании постоянства
внутренней среды и обеспечении интеграции функций автономной, эндокринной,
соматической систем. Этот небольшой по объему, но важный по функциям отдел
лежит на дне и по бокам III желудочка, вентральное таламуса. Он включает в себя
такие анатомические структуры, как серый бугор, воронку, которая
заканчивается
|
|
|
Рис. 3.27 Расположение основных ядерных групп гипоталамуса 1 —
паравентрикулярное ядро, 2 —
спайка, 3 — свод, 4 —
мозолистое тело, 5 — таламус, 6 —
шишковидное тело, 7 — водопровод, 8
— средний мозг, в —
заднее ядро, 10 — сосцевидное
тело, 11 — дорсомедиальное
ядро, 12 — латеральное ядро, 13 — вентромедиальное ядро, 14 — нейрогипофиз, 16 — аденогипофиз, 16 — зрительный перекрест, 17 — супраоптическое ядро. |
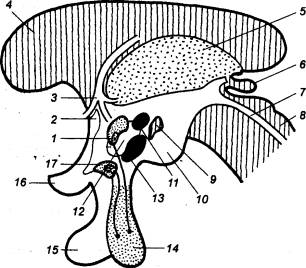
гипофизом, и сосцевидные, или
мамиллярные, тела. Верхнюю границу гипоталамуса формируют конечная
пластинка и зрительный перекрест (рис.
3.27). Сбоку гипоталамус ограничен зрительным трактом и внутренней
капсулой, а сзади примыкает к среднему мозгу.
В нейрональной сети гипоталамуса
можно выделить несколько десятков ядер, которые топографически подразделяются
на пять групп. Предоптическая группа, наиболее близкая к конечному мозгу,
состоит из перивентрикулярного, медиального и латерального
предоптических ядер. Передняя группа включает супрахиазматическое,
супраоптическое и паравентрикулярное ядра. Отростки нейронов двух последних
четко отграниченных ядер идут к задней доле гипофиза — нейрогипофизу. Средняя
группа ядер гипоталамуса образована вентромедиальным и дорсомедиальным
ядрами. В состав наружной группы
входят латеральное гипоталамическое и серобугорное ядра. И
наконец, задняя группа сформирована из заднего гипоталамического ядра,
перифорникального ядра и нескольких ядер сосцевидного тела.
Большинство ядер гипоталамуса имеют
плохо очерченные границы и, за малым исключением (супраоптическое и
паравентрикулярное ядра), их трудно рассматривать как центры с узкой
локализацией специфических функций. Более перспективным является деление гипоталамуса
на области и зоны, обладающие определенной функциональной спецификой. Так,
например, некоторые ядра предоптической и передней групп объединяются в гипофизотропную
область, нейроны которой продуцируют так называемые рилизинг—факторы,
(либерины) и ингибирующие факторы (статины), регулирующие
деятельность передней доли гипофиза — аденогипофиза.
Срединная группа ядер формирует медиальный
гипоталамус, где имеются своеобразные нейроны—датчики, реагирующие на
различные изменения внутренней среды организма (температуру крови,
водно—электролитный состав плазмы, содержание гормонов в крови). Посредством
нервных и гуморальных механизмов медиальный гипоталамус управляет деятельностью
гипофиза.
Латеральный гипоталамус представляет собой безъядерную
зону, в которой сосредоточены проводниковые элементы (медиальный пучок
переднего мозга), формирующие пути к верхним и нижним отделам стволовой части
мозга. Нервные клетки в латеральном гипоталамусе расположены диффузно. В целом
гипоталамус как важный интегративный центр имеет богатые афферентные и
эфферентные связи с различными отделами мозга.
Через систему волокон дорсального
продольного пучка (Шютца) гипоталамус получает афферентный вход от
ретикулярной формации среднего мозга. Параллельно этому каналу афферентные
импульсы от среднего мозга могут поступать в гипоталамус по сосцевидно—покрышечному
пучку. Гипоталамус обладает двусторонними связями с таламусом и через
передние таламические ядра — с корой больших полушарий. Афферентный вход от
лимбической системы мозга осуществляется через медиальный пучок конечного
мозга, проходящий в латеральном гипоталамусе.
Паллидо—гипоталамический путь
связывает гипоталамус с базальными ганглиями переднего мозга. Кроме того,
афферентные входы гипоталамуса дополняются непрямыми мозжечково—гипоталамическими
и ваго—супраоптическими связями. Наличие последних подтверждается и
электрофизиологическими данными о представительстве в гипоталамусе висцеральной
афферентации, приходящей по чувствительным волокнам блуждающего и чревных
нервов.
Основные эфферентные связи
гипоталамуса с вегетативными и соматическими ядрами головного и спинного мозга
осуществляются через полисинаптические пути ретикулярной формации ствола.
Эти пути дополняются короткими
эфферентными путями, идущими в таламус и через его посредство далее в кору
больших полушарий. В системе мозгового свода проходят эфферентные волокна от
гипоталамуса в лимбическую кору. Через сосцевидно—покрышечный пучок эфферентная
импульсация от гипоталамуса может достигать среднего мозга. Особо выделяется
мощный пучок нисходящих волокон, идущих от супраоптического ядра гипоталамуса в
нейрогипофиз.
Таким образом, сама структурная
организация гипоталамуса свидетельствует о функциональной значимости этого
отдела головного мозга. Вместе с тем эта организация настолько сложна, что в
вопросе об основных принципах ее построения еще остается много загадочного.
3.7.4. Роль гипоталамуса в регуляции вегетативных функций
Исследование вегетативных и
соматических реакций, вызванных локальным электрическим раздражением различных
областей гипоталамуса, позволило В. Гессу (1954) выделить в этом отделе мозга две
функционально дифференцированные зоны. Раздражение одной из них — задней
и латеральной областей гипоталамуса — вызывает типичные симпатические
эффекты', расширение зрачков, подъем кровяного давления, увеличение частоты
сердечных сокращений, прекращение перистальтики кишечника и т. д. Разрушение
данной зоны, напротив, приводило к длительному снижению тонуса симпатической
нервной системы и контрастному изменению всех перечисленных выше показателей.
Гесс назвал область заднего гипоталамуса эрготропной и допустил, что
здесь локализованы высшие центры симпатической нервной системы.
Другая зона, охватывающая предоптическую
и переднюю области гипоталамуса, получила название трофотропной, так
как при ее раздражении наблюдались все признаки общего возбуждения
парасимпатической нервной системы, сопровождавшиеся реакциями,
направленными на восстановление и сохранение резервов организма.
Однако дальнейшие исследования
показали, что гипотеза о наличии двух антагонистических зон гипоталамуса
является слишком обобщенной и не может объяснить различные функции этого Отдела
мозга. Так были получены факты, противоречащие узкой локализации симпатических
и парасимпатических центров в гипоталамусе. Оказалось, что сосудосуживающий
эффект может быть получен при раздражении и задней, и передней областей
гипоталамуса и, следовательно, симпатические нейроны присутствуют в этих обеих
областях. Равным образом клетки, которые активируют постганглионарные
холинергические сосудорасширяющие нейроны, тоже довольно широко распространены
в пределах гипоталамуса (см. разд. 5.5.3).
Детальное исследование изменений в
сердечно—сосудистой системе показало, что локальное раздражение определенных
отделов гипоталамуса сопровождается диаметрально противоположными изменениями
кровотока в «ядре» и «оболочке» тела. Например, при увеличении кровотока в
скелетных мышцах может наблюдаться его снижение в сосудах кожи и органах
брюшной полости. При раздражении гипоталамуса возможны не только вегетативные
реакции, но и целый ряд соматических эффектов, таких как изменение позы или
увеличение частоты дыхания.
Эти факты свидетельствуют о том, что
гипоталамус вряд ли является статическим и локальным объединением эрготропной и
трофотропной зон. Предпочтительнее представлять себе гипоталамус как важный интегративный
центр автономных, соматических и эндокринных функций, который отвечает за
реализацию сложных гомеостатических реакций и входит в иерархически
организованную систему отделов головного мозга, регулирующих висцеральные
функции.
Системность гомеостатических реакций
можно продемонстрировать опять—таки на примере регуляции гемодинамики.
Известно, что саморегуляция сосудистого тонуса осуществляется за счет функции
сосудистого центра продолговатого мозга, который работает как следящая
система. Этот уровень интеграции достаточен для осуществления простых
рефлекторных реакций в ответ на информацию от баро— и механорецепторов
сосудистого русла. Вместе с тем более сложные вазомоторные реакции, связанные,
например, с терморегуляцией или локомоторными актами, реализуются с участием
гипоталамуса, который связан с сосудистым центром и с сосудодвигательными
нейронами спинного мозга.
В то же время сам гипоталамус имеет
связи с корой головного мозга, представляющей еще более высокий уровень
интеграции. Такие связи обнаружены, например, между корой и латеральной
областью гипоталамуса, отвечающей за приспособительные реакции сосудистой
системы при физической нагрузке. Очевидно, по этим путям распространяются
модулирующие влияния коры на деятельность гипоталамуса.
Таким образом, в регуляцию
вегетативных реакций вовлекается целая система центров, представленных на всех
уровнях головного мозга. Гипоталамус является одним из уровней данной системы,
и это во многом определяет сложность и адаптивный характер регулируемых им
вегетативных реакций.
3.7.5. Терморегуляционная функция
гипоталамуса
Целый ряд экспериментальных и
клинических данных указывает на то, что гипоталамус является интегративным
центром терморегуляции (см. разд. 10.4.5). Так, например, перерезка
стволовой части мозга у кошки на уровне верхней границы среднего мозга нарушает
терморегуляцию и превращает теплокровное животное в холоднокровное. Локальное раздражение
задних отделов гипоталамуса показало, что здесь находятся структуры,
регулирующие процесс теплопродукции. Эффект от раздражения проявляется в
интенсификации обменных процессов, увеличении частоты сердечных сокращений и
появлении дрожания скелетной мускулатуры. Разрушение задних отделов
гипоталамуса вызывает подавление теплопродукции и снижение температуры тела.
Передние отделы гипоталамуса, в
частности паравентрикулярные ядра, ответственны за интеграцию процессов
теплоотдачи. Раздражение этой области вызывает расширение кожных сосудов,
усиление потоотделения» интенсификацию дыхания. Повреждение переднего
гипоталамуса сопровождается нарушением теплоотдачи и повышением температуры
организма — гипертермией. При регистрации электрической активности одиночных
клеток в переднем гипоталамусе были обнаружены так называемые тепловые
нейроны, у которых локальное нагревание вызывает увеличение частоты
импульсной активности. Как правило, усиление разрядов этих нейронов
предшествует комплексу реакций, увеличивающих теплоотдачу.
Тепловые нейроны переднего
гипоталамуса могут быть отнесены к категории внутренних терморецепторов,
которые в естественных условиях реагируют на температуру крови и вместе с
кожными терморецепторами рефлекторно возбуждают эфферентные нейроны
гипоталамуса, контролирующие теплоотдачу на периферии.
3.7.6. Участие гипоталамуса в регуляции поведенческих
реакций
Локальные электрические раздражения
определенных зон гипоталамуса могут вызывать направленные на выживание особи
поведенческие комплексы, которые включают моторные, вегетативные и
гормональные компоненты. Так, в заднем гипоталамусе обнаружена область,
электрическая стимуляция которой вызывает комплекс реакций, характерных для
пищевого поведения: поиск пищи, обильное слюноотделение, усиленная моторика и
кровоснабжение кишки, снижение мышечного кровотока.
Показано, что повреждение небольшого
участка в латеральном гипоталамусе ведет к полному отказу от пищи (афагия)
и воды (адипсия), истощению и гибели животного. Эта зона названа «зоной
голода».
Разрушение вентромедиальных ядер
гипоталамуса, наоборот, вызывает чрезмерное потребление пищи (гиперфагию)
и ожирение. Здесь локализован «центр насыщения», нейроны которого, по всей вероятности,
обладают прямой химической чувствительностью к составу крови, зависимому от
периодичности питания. Нейроны центра насыщения по мере потребления пищи
активируются и через внутригипоталамические тормозные связи подавляют функции
центра голода, что» как следствие, вызывает устранение комплекса реакций,
связанных с пищевым поведением.
Эта привлекательная гипотеза в целом
еще не получила экспериментального подтверждения, так как тормозные
взаимодействия между центрами не идентифицированы, да и само понятие центров
голода и насыщения является скорее абстрактным, нежели соотнесенным с
представлением о четко очерченных анатомических структурах.
Дело в том, что области
гипоталамуса, связанные с конкретными поведенческими реакциями, широко
перекрываются и зачастую их удаление вызывает содружественные нарушения общего
поведения животных. Например, разрушение латерального гипоталамуса помимо
афагии ведет еще и к снижению двигательной активности животных, угнетению
эмоций, ослаблению устойчивости к стрессу. Эти факты объясняются тем, что при
повреждении латерального гипоталамуса нарушаются функции не только одной
изолированной системы. Как известно, через латеральный гипоталамус проходят
волокна мощного медиального пучка конечного мозга, который связывает между
собой базальные отделы переднего мозга, гипоталамус и покрышку среднего мозга.
Сюда же от стволовой части мозга подходят восходящие дофаминергические и
норадренергические пути, поражение которых вызывает комплекс нарушений, сходных
с теми, которые возникают при повреждении латерального гипоталамуса.
Таким образом, латеральный
гипоталамус является общим путем, в котором сходятся воздействия разнообразных
интегративных систем мозга, и нет ничего удивительного в том, что его
повреждение вызывает комплексные нарушения поведенческих реакций.
В связи с этим следует отметить, что
в медиальном пучке конечного мозга, так же как и в заднем гипоталамусе,
обнаружены зоны, связанные с регуляцией полового поведения. Показано,
что крысы с хронически вживленными в эти зоны электродами довольно быстро
обучались нажимать на рычаг, который включает электрическое раздражение их
собственного мозга. Такое производимое по собственному почину раздражение
действует как «вознаграждение». Если животным предоставлялся свободный выбор,
то голодные крысы гораздо быстрее устремлялись к рычагу самостимуляции, чем к
лежащей рядом пище. Максимальная частота самораздражения достигала 5000 нажатий
рычага за 1 ч.
Естественно, что о субъективных
ощущениях животных мы ничего не знаем, и вопрос о возможности использовать
эксперименты на животных для объяснения человеческих эмоций долгое время
оставался спорным. Однако работы, проведенные в условиях нейрохирургической
клиники, показали, что раздражение аналогичных участков мозга у людей вызывало
чувства радости, удовлетворения, сопровождавшиеся эротическими переживаниями.
Очевидно, эти положительные эмоции являются компонентами полового поведения,
которое регулируется задним гипоталамусом. Вместе с тем судить о локализации
здесь «центра удовольствия» несколько опрометчиво, так как система регуляции
полового поведения включает и другие отделы головного мозга, в том числе
лимбическую кору.
Материалы клинических данных
свидетельствуют о том, что гипоталамус определяет правильную периодичность
функций, связанных с размножением. Опухолевые процессы в области
гипоталамуса могут вызывать быстрое половое созревание, нарушение
менструального цикла, половую слабость и ряд других дисфункций.
Как показали опыты с локальным
раздражением, гипоталамус участвует в регуляции агрессивного поведения
животных. При раздражении передних отделов гипоталамуса кошка принимает
угрожающую позу, оскаливает зубы, шипит и выпускает когти. Эта реакция
сопровождается вегетативными компонентами — увеличением частоты сердечных
сокращений и пиломоторным эффектом — возбуждением мышц, поднимающих волосы. В
связи с тем, что данная реакция не имеет объекта агрессии, она называется ложной
яростью.
Иная картина наблюдается при
раздражении некоторых латеральных отделов гипоталамуса. В данном случае
агрессия имеет четкую направленность против контрольного животного, без лишних
движений и агрессивных демонстраций. Эти факты говорят о том, что каждая форма
поведения имеет свою эмоциональную окраску, в создании которой гипоталамус как
интегративный центр играет существенную роль.
Как регуляторный орган гипоталамус
принимает участие в чередовании состояний сна и бодрствования. В клинике
при повреждении гипоталамуса описаны случаи перехода в состояние летаргического
сна — обездвиженность, понижение обмена веществ, ослабление реакций на
внешние раздражители. Сноподобное состояние у животных можно вызвать при
электрическом раздражении некоторых зон медиального гипоталамуса. Напротив,
задний гипоталамус имеет решающее значение в поддержании состояния бодрствования.
Переход от сна к бодрствованию и обратно сопровождается изменением соматических
(мышечный тонус) и вегетативных (частота сердечных сокращений, перистальтика
кишки) процессов, интеграция которых осуществляется гипоталамусом.
В процессы регуляции гипоталамусом
приспособительных поведенческих реакций входит и его участие в поддержании
водного баланса организма. Отсутствие воды создает мощное побуждение
(мотивацию), направленное на устранение дефицита жидкости. В создании этой
мотивации, в появлении чувства жажды участвует гипоталамус, в передней области
которого обнаружены нейроны с осморецепторной функцией. Эти нейроны
возбуждаются при изменении осмотического давления крови и запускают целый
комплекс соматических и эндокринных реакций, направленных на устранение данной
мотивации.
Даже это краткое перечисление
регуляторных функций гипоталамуса свидетельствует о его важной роли в
организации поведения у животных.
Вместе с тем принцип нейронной
организации гипоталамуса до сих пор мало исследован и остается загадкой, как
такое маленькое образование способно управлять многими системными реакциями
организма. Вероятно, исследование структуры и функции нейронных цепей
гипоталамуса обеспечит дальнейший прогресс в нашем познавании этого отдела
мозга.
3.7.7. Гипоталамо—гипофизарная система
Одной из важнейших функций
гипоталамуса является регуляция деятельности гипофиза (см. разд. 6.2). Гипофиз
— главная железа внутренней секреции — состоит из передней, задней и
промежуточной долей. В процессе эмбриогенеза передняя доля, или аденогипофиз,
развивается из выпячивания глотки, а задняя, или нейрогипофиз,
формируется из промежуточного мозга. Как передняя, так и задняя доли гипофиза
находятся под контролем гипоталамуса, однако механизмы их регуляции различны.
Нейрогипофиз является органом,
депонирующим два гормона: антидиуретический (АДГ, вазопрессин) и окситоцин.
Как уже упоминалось, местом синтеза этих гормонов являются супраоптическое и
паравентрикулярное ядра гипоталамуса. Формируясь в нейросекреторных клетках
этих ядер, гормоны в виде гранул транспортируются по их аксонам и после
разрушения гранул выделяются в капиллярную сеть нейрогипофиза.
Антидиуретический гормон регулирует обратное всасывание воды в почечных
канальцах и воздействует на гладкую мускулатуру артериол, повышая таким образом
артериальное давление. Окситоцин стимулирует сокращение гладкой мускулатуры
матки и молочных желез.
Регуляция секреции обоих гормонов
осуществляется по механизму нейрогуморального рефлекса, афферентное
звено которого представлено нервными путями от осморецепторов или
механорецепторов до гипоталамуса, а эфферентное звено — поступающим в кровяное
русло гормоном. Кроме того, регуляция функций молочной железы может
осуществляться на первых порах чисто гуморальным путем за счет изменения гормонального
фона во время беременности.
Если гормоны задней доли гипофиза
продуцируются нейросекреторными клетками гипоталамических ядер, то все гормоны
передней доли секретируются клетками самого аденогипофиза. В зависимости
от мишеней, на которые направлено их действие, гормоны аденогипофиза
подразделяются на гландотропные, влияющие на другие периферические
эндокринные железы, и эффекторные, воздействующие непосредственно на
ткани. К первой группе относятся кортикотропин (адренокортикотропный
гормон, АКТГ) — регулирует секрецию глюкокортикоидов в коре надпочечников; тиреотропный
гормон (ТТГ) — стимулирует синтез и секрецию йодсодержащих гормонов
щитовидной железы; гонадотропные гормоны — фолликулостимулирующий (ФСГ)
и лютеинизирующий (ЛГ) — регулируют деятельность половых желез.
Эффекторные гормоны аденогипофиза
представлены соматотропным гормоном (СТГ, гормон роста), регулирующим
рост костей, и пролактином, который стимулирует рост молочных желез и секрецию
молока. К числу эффекторных гормонов относятся также меланоцитстимулирующий
гормон промежуточной доли гипофиза (МСГ, интермедии), являющийся
регулятором кожной пигментации.
В 70—е гг. было установлено, что
секреторная функция аденогипофиза находится под контролем гипоталамуса. Этот
контроль осуществляется нейрогуморальным путем за счет гормонов, выделяемых
гипофизотропной зоной гипоталамуса (в медиальном гипоталамусе). Оказалось, что
секреция всех известных гормонов аденогипофиза регулируется гипофизотропными
гормонами гипоталамуса, которые являются как бы гормонами гормонов.
Гипофизотропные гормоны представляют собой пептиды с низкой молекулярной массой
(около 4 кДа) и подразделяются на стимулирующие, или так называемые рилизинг—факторы
(либерины), и тормозящие секрецию ингибирующие факторы (статины),
Высвобождаясь из нервных окончаний,
гипофизотропные гормоны через сосуды гипоталамо—гипофизарной портальной системы
попадают в аденогипофиз и там воздействуют на клетки, секретирующие тот или
иной тропный гормон. В 1975 г. из гипоталамуса и гипофиза была выделена еще
одна группа пептидов — энкефалины и эндорфины, которые оказывают
на нервные клетки морфиноподобное действие и, по—видимому, играют существенную
роль в регуляции вегетативных процессов и поведения.
Секреция гипофизотропных гормонов
гипоталамуса регулируется по принципу отрицательной обратной связи.
Установлено, что при повышении содержания в плазме крови гормонов
периферических желез внутренней секреции уменьшается поступление соответствущих
рилизинг—факторов в портальную гипоталамо—гипофизарную систему и тем самым
снижается секреция того или иного тройного гормона аденогипофиза. Параллельно с
этим механизмом деятельность гипоталамо—гипофизарной системы может
регулироваться и за счет нервных влияний, приходящих от лимбической системы и
среднего мозга через латеральный гипоталамус. Так, известно, что при сильных
болевых или других стрессорных воздействиях у животных наблюдается усиленное
выделение кортикотропина и, наоборот, снижение секреции гонадотропных гормонов.
Механизм этой реакции обусловлен изменениями секреции соответствующих
рилизинг—факторов, которые вызываются импульсацией, приходящей в
гипофизотропную зону гипоталамуса из лимбической коры и среднего мозга.
Существует точка зрения, что сигналы от этих центров поступают через дофамин— и
норадренергические пути.
Таким образом, за счет нервных
влияний секреция аденогипофиза приводится в соответствие с мотивацией и
биологической направленностью реакций организма.
Гипоталамус как нервный центр,
участвующий в формировании биологических побуждений к действию, или мотиваций,
тесно связан с лимбической системой мозга. Под лимбической системой
понимают морфофункциональное объединение, которое включает в себя
филогенетически старые отделы коры переднего мозга, а также ряд подкорковых
структур, которые регулируют функции внутренних органов, обусловливающих
эмоциональную окраску поведения и его соответствие имеющемуся субъективному
опыту (см. разд. 5.5).
3.8.1. Анатомические структуры лимбической системы
В состав лимбической коры входит древняя
кора (палеокортекс), формирующая обонятельный мозг и состоящая из
обонятельных луковиц, обонятельного бугорка, прозрачной перегородки и
прилежащих областей коры (препириформная, периамигдалярная и диагональная области).
Следующий компонент лимбической системы — старая кора (археокортекс),
объединяющая сместившийся в процессе эволюции к височной доле гиппокамп,
зубчатую извилину, основание гиппокампа (субикулум) и расположенную над
мозолистым телом поясную извилину (рис. 3.28).
Древняя и старая кора, которые
обозначают как аллокортекс, граничат с пятислойной межуточной корой, или
мезокортексом, переходящим непосредственно в шестислойную новую кору
— неокортекс, или изокортекс. Мезокортекс формируется прилежащей к
древней коре островковой долей и граничащей со старой корой парагиппокампальной
извилиной (энториальная область и предоснование гиппокампа, или пресубикулум),
которые также включены в состав лимбической системы.
Из подкорковых структур в
лимбическую систему входят расположенные в медиальной стенке височной доли миндалевидное
тело и ядра перегородки конечного мозга. Многие исследователи
причисляют к лимбической системе также переднее таламическое ядро, сосцевидные
тела и гипоталамус.
Все многочисленные формирования
лимбической коры кольцеобразно охватывают основание переднего мозга и являются
своеобразной границей между новой корой и стволовой частью мозга.
Лимбическая система характеризуется
обилием двусторонних связей с другими отделами мозга и внутри самой системы.
Так, например, установлено наличие мощных связей лимбической системы с
гипоталамусом.
Через гипоталамус и сосцевидные тела
лимбическая система соединена с центральным серым веществом и ретикулярной
формацией среднего мозга.
|
|
|
Рис. 3.28 Расположение лимбической системы в виде кольца по краю
неокортекса (А); афферентные и эфферентные связи лимбической системы (Б) 1 —
поясная извилина, 2 — мозолистое тело, 3 — центральная борозда, 4 — теменная
доля, 5 — шпорная борозда, 6 — затылочная доля, 7 — гиппокамп, 8 —
парагиппокампальная извилина, 9 — височная доля, 10 — крючок, 11 —
миндалевидное тело, I2 — обонятельная луковица, 13 —
передняя спайка, 14 — лобная доля, 16 — передний таламус, 16 — пояс, 17 —
свод, 18 — терминальная полоска, 10 — сосцевидное тело, 20 — гипоталамус, 21
— медиальный пучок конечного мозга,
22 — перегородка. |
К миндалине и гиппокампу идут пути
от височной доли коры, передающие информацию от зрительной, слуховой и
соматической сенсорных систем. Установлены связи лимбической системы с лобными
долями коры переднего мозга. Наконец, в пределах лимбической системы
идентифицированы сложные циклические связи, создающие условия для циркуляции
возбуждения по сложным круговым путям. Примером такой циклической связи может
служить так называемый круг Папеса (Papez), идущий от гиппокампа через свод —
сосцевидное тело — переднее ядро таламуса — кору поясной извилины и
пресубикулум обратно к гиппокампу.
Очевидно, сложность связей и
внутренней организации лимбической системы свидетельствует об ее участии в
интеграции функций новой коры и стволовых образований головного мозга.
3.8.2. Функции лимбической системы
Исследование функции лимбической
системы у высших позвоночных началось сравнительно недавно. Длительное время в
физиологии существовала точка зрения, что аллокортекс имеет только обонятельную
функцию, и это нашло выражение в термине обонятельный мозг. Вместе с тем
экспериментальные данные показывают, что помимо обонятельных афферентов в
аллокортексе обнаруживаются проекции и других афферентных систем,
конвергирующих к одним и тем же нервным структурам. Это и ряд других факторов
заставили отказаться от традиционных взглядов и расширить наши представления о
функциях древней и старой коры.
В 1937 г. американский невропатолог Дж.
Папес выдвинул гипотезу, согласно которой данные структуры мозга образуют
единую систему (круг Папеса), ответственную за осуществление врожденных
поведенческих актов и формирование эмоций. В 1952 г. другой американский
исследователь, П. Маклин, развивая предположения Папеса, ввел понятие лимбическая
система, представляя этим термином сложную функциональную систему,
обеспечивающую постоянство внутренней среды и контроль видоспецифических
реакций, направленных на сохранение вида. Теоретические и практические
разработки Папеса и Маклина послужили мощным толчком для дальнейших
исследований в этой области.
Эти исследования показали, что
локальное раздражение различных отделов лимбической системы вызывает
разнообразные вегетативные эффекты и влияет на деятельность внутренних органов
(см. разд. 5.5.4). Так, раздражение ядер миндалевидного комплекса приводит к
изменениям частоты сердечного ритма, дыхательных движений, сосудистого тонуса.
В ряде случаев раздражение миндалин влияет на деятельность пищеварительного тракта,
изменяя перистальтику тонкой кишки, стимулируя секрецию слюны, произвольное
жевание и глотание. Описано влияние миндалин на сокращения мочевого пузыря,
матки, пилоэрекцию и сокращение третьего века. Все эти разнообразные реакции
могут иметь различный знак и характеризоваться активацией или угнетением
висцеральных функций.
Существуют указания на то, что
кортикальные и медиальные ядра миндалевидного комплекса имеют отношение к
регуляции пищевого поведения. Однако большей частью попытки связать тип реакции
с определенным ядром миндалевидного комплекса не дали четких результатов. Факты
указывают скорее на то, что функциональное представительство в миндалевидном
комплексе, да и в других структурах
лимбической системы (например, в гиппокампе)' не дифференцировано. В некоторых
случаях негативные результаты объясняются генерализованным, распространяющимся
на соседние участки нервной ткани действием электрических стимулов.
Вместе с тем характер вегетативных
реакций, вызванных раздражением различных отделов миндалины, идентичен эффектам
при электрической стимуляции гипоталамуса, и это позволяет думать, что регулирующие
влияния лимбической системы опосредованы нижележащими вегетативными центрами.
Скорее всего, изменяя в ту или иную
сторону возбудимость гипоталамических центров, лимбическая система определяет
знак соответствующей вегетативной реакции. Так формируется многоэтажная,
построенная по иерархическому принципу система управления вегетативной сферой,
интегрирующая вегетативные и соматические реакции.
Как известно, эмоциональная окраска
поведенческих реакций определяется не только вегетативными компонентами, но и
соответствующими эндокринными сдвигами. В этом плане представляют интерес
данные о влиянии лимбической системы на деятельность желез внутренней секреции.
Установлено, что длительное (60 мин) раздражение ядер миндалевидного комплекса
у обезьян вызывает повышение содержания кортикостероидов в плазме крови.
Низкочастотная (12—36 имп/с) стимуляция гиппокампа, напротив, уменьшает
содержание кортикостероидов, которые выделяются надпочечниками при нанесении
стрессорных раздражителей. Очевидно, нисходящие влияния этих структур
лимбической системы на гипоталамус, а через него на гипофиз изменяют продукцию
кортикотропина, который регулирует секрецию кортикостероидов.
Таким образом, изменяя гормональный
фон, лимбическая система в естественных условиях может участвовать в
формировании побуждений к действию (мотиваций) и регулировать реализацию самих
действий, направленных на устранение побуждения, усиливая или ослабляя
эмоциональные факторы поведения.
3.8.3. Роль лимбической системы в
формировании эмоций
Сведения об участии различных
отделов лимбической системы в формировании эмоций неоднозначны. Наибольшее их
количество относится к роли миндалевидного комплекса и поясной извилины в этих
процессах. При локальном электрическом раздражении ядер миндалевидного
комплекса могут быть получены эмоциональные реакции типа страха, гнева, ярости
и агрессии. Двустороннее удаление височных долей вместе с миндалиной и гиппокампом
вызывает у макак—резусов целый ряд сдвигов в эмоциональной сфере. Как правило,
агрессивные обезьяны после этой операции становятся спокойными и доверчивыми. У
животных наблюдается гиперорализм, когда все незнакомые предметы без
разбора заталкиваются в рот.
Удаление
височных долей вызывает у обезьян гиперсексуальность, причем их половая
активность может быть направлена даже на неодушевленные предметы. Наконец,
послеоперационный синдром сопровождается так называемой психической
слепотой. Животные утрачивают способность правильной оценки зрительной и
слуховой информации, и эта информация никак не связывается с собственным
эмоциональным настроем обезьян. Так, макаки без разбора исследуют все, даже
опасные для себя предметы. Возникновение психической слепоты связывают с
послеоперационным нарушением передачи сенсорной информации от височной доли к
гипоталамусу. Эта точка зрения подкрепляется электрофизиологическими данными,
свидетельствующими о том, что различные сенсорные раздражения изменяют частоту
импульсной активности нейронов миндалевидного комплекса, который, очевидно,
участвует в оценке поступающей из внешней среды информации.
Согласно теории Папеса, сенсорные
пути на уровне таламуса расходятся, причем один путь идет в проекционные зоны
коры, где обеспечивается восприятие, а второй — в лимбическую систему.
Вероятно, в этой системе происходит
оценка поступающей информации, ее сопоставление с субъективным опытом и запуск
соответствующих эмоциональных реакций через гипоталамические структуры.
Информация об аффективном состоянии организма может поступать от стволовых
структур в неокортекс либо прямо от миндалевидного комплекса, либо через
поясную извилину, которая связана с лобными, теменными и височными долями коры
полушарий большого мозга. Вероятно, эти пути связаны с восприятием
эмоционального фона, однако детальные механизмы этого восприятия еще нуждаются
в разработке.
Как уже отмечалось, лимбическая
система принимает участие в запуске тех эмоциональных реакций, которые уже
апробированы в ходе жизненного опыта. В этом плане привлекают внимание
исследования, констатирующие участие лимбической системы в процессах
сохранения памяти. Так, например, удаление гиппокампа (аммонова
рога) (От греч. hippokampos — мифическое чудовище с туловищем коня и рыбьим хвостом.) вызывает
у людей полное выпадение памяти на недавние события. Электрическое раздражение
парагиппокампальной извилины во время нейрохирургических операций может
сопровождаться появлением мимолетных воспоминаний. Двустороннее удаление
гиппокампа у обезьян и крыс приводит к нарушению способности выполнять ту или
иную последовательность поведенческих актов. Описанные факты привели к
заключению, что гиппокамп играет определенную роль в процессах памяти, и
послужили толчком для дальнейших исследований этой структуры.
Оказалось, что для строения
гиппокампа характерно наличие четко отграниченных слоев с преимущественным
расположением в них либо тел, либо отростков нервных клеток. Нейронные цепи
гиппокампа представляют собой стереотипные микросети, которые состоят из
возбуждающих волокон энториальной коры, клеток—зерен и пирамидных клеток с
аксонами, идущими через свод к перегородке. Аксоны пирамидных клеток образуют
коллатерали, направляющиеся к нейронам других частей гиппокампа.
Такая стереотипная слоистая
структура гиппокампа делает его очень удобным объектом для изучения функции
нейронных сетей на перфузируемых срезах мозга. Электрофизиологические
исследования срезов и тотальных препаратов гиппокампа показали, что для этой
структуры характерны периодически низкочастотные (4—5 в 1 с) электрические
колебания — τ (тета)—ритм. Эти колебания сопряжены с правильным
чередованием возбуждающих и тормозных постсинаптических потенциалов пирамидных
клеток гиппокампа и, вероятно, отражают синхронизацию деятельности его нейронных
элементов.
Способность генерировать ритмическую
активность, по всей видимости, зависит от упорядоченной слоистой структуры
гиппокампа, которая создает условия для циркулирования возбуждения по нейронным
цепям, лежащего в основе одного из нейронных механизмов памяти.
Об участии гиппокампа в процессах консолидации
памяти свидетельствуют также изменения свойств его нейрональных синапсов
после тетанической стимуляции. При высокочастотной стимуляции энториальной
области коры кролика наблюдается длительное, достигающее нескольких часов, а
иногда даже дней, возрастание амплитуды синаптических потенциалов клеток—зерен
гиппокампа. В дальнейшем при микроэлектродном исследовании толстых срезов
гиппокампа было установлено, что изменение эффективности синаптического проведения
после тетанической стимуляции обусловлено повышением секреции медиатора из
пресинаптических окончаний. Параллельные электронно—микроскопические
исследования показали, что после тетанической стимуляции наблюдалось увеличение
числа шипиков на дендритах нейронов гиппокампа.
Эти факты убеждают в том, что
пластичность нейронных цепей гиппокампа является предпосылкой его участия в
формировании нейронных механизмов памяти. Однако было бы упрощением считать
гиппокамп единственным хранилищем следов памяти у позвоночных. Пластичность
является весьма распространенным свойством нейронов, и поэтому большинство
исследователей склоняются к тому, что функция памяти не является прерогативой
какой—либо одной структуры, а обусловлена содружественными действиями многих центров
головного мозга. Существенным звеном в этой системе являются связи гиппокампа с
неокортексом.
Функциональная роль этих связей
подтверждается физиологическими экспериментами. При одновременной регистрации
электрической активности гиппокампа и неокортекса наблюдаются реципрокные
взаимоотношения между ними. Когда в гиппокампе возникает медленноволновый
Θ—ритм, в неокортексе доминирует высокочастотная низкоамплитудная
активность и, наоборот, медленноволновой активности энцефалограммы соответствует
высокочастотная активность гиппокампа.
Наиболее выраженное усиление
Θ—ритма в гиппокампе обнаруживается на начальных стадиях выработки
условного рефлекса и сочетается с состоянием настороженности и сосредоточения
внимания при формировании ориентировочной реакции.
Удаление гиппокампа у животных
нарушает процессы внутреннего торможения и снижает способность к угашению
потерявших свое адаптивное значение условнорефлекторных реакций. Одновременно
затрудняется упрочение условного рефлекса в связи с резким усилением ориентировочной
реакции. Следовательно, гиппокамп, как, впрочем, и другие структуры лимбической
системы, существенно влияет на функции неокортекса и на процессы научения. Это
влияние осуществляется в первую очередь за счет создания эмоционального фона,
который в значительной степени отражается на скорости образования любого
условного рефлекса.
Таким образом, лимбическую систему
как одно из наиболее древних образований мозга нельзя считать простым
атавизмом. Это важный отдел головного мозга, функционально связанный с
неокортексом и стволовыми структурами, образующими вместе систему
координации висцеральных и соматических функций организма.
3.9.
БАЗАЛЬНЫЕ ЯДРА И ИХ ФУНКЦИИ
Базальные ядра, названные гистологами прошлого века
ганглиями, представляют собой структуры ядерного типа, которые располагаются в
толще белого вещества переднего мозга ближе к его основанию. У млекопитающих к
базальным ядрам относятся сильно вытянутое в длину и изогнутое хвостатое
ядро и заложенное в толще белого вещества чечевицеобразное ядро. Двумя
белыми пластинками оно подразделяется на три части: наиболее крупную, лежащую
латерально скорлупу, и бледный шар, состоящий из внутреннего и
внешнего отделов (рис. 3.29).
|
|
|
Рис. 3.29 Афферентные и эфферентные связи базальных
ядер 1 — хвостатое ядро, 2 —
скорлупа, 3 — черное вещество, 4 —
таламус, 5, 5* — внутренний и
внешний отделы бледного шара, 6 — субталамическое
ядро, 7 — сенсомоторная кора,
8 — двигательная кора, 9 — пути к стволовой части мозга;
стрелками обозначены афферентные пути, эфферентные пути, межъядерные
взаимодействия. |
Эти анатомические образования
формируют так называемую стриопаллидарную систему (От лат. striatus — полосатый и pallidus — бледный.), которая по филогенетическим
и функциональным критериям разделяется на древнюю часть палеостриатум и новую —
неостриатум. Палеостриатум представлен бледным шаром, а неостриатум,
появляющийся впервые у рептилий, состоит из хвостатого ядра и скорлупы, которые
объединяются под названием полосатого тела, или стриатума. Хвостатое
ядро и скорлупа связаны анатомически и характеризуются чередованием белого и
серого вещества, что оправдывает возникновение термина полосатое тело.
К стриопаллидарной системе часто
относят также субталамическое ядро (люисово тело) и черное вещество
среднего мозга, которые образуют с базальными ядрами функциональное единство.
Полосатое тело состоит главным образом из мелких клеток, аксоны которых
направляются к бледному шару и черному веществу среднего мозга.
Полосатое тело является своеобразным
коллектором афферентных входов, идущих к базальным ядрам. Главными источниками
этих входов служат новая кора (преимущественно сенсомоторная), неспецифические
ядра таламуса и дофаминергические пути от черного вещества.
В противоположность полосатому телу бледный
шар состоит из крупных нейронов и является сосредоточением выходных,
эфферентных путей стриопаллидарной системы. Аксоны локализованных в бледном
шаре нейронов подходят к различным ядрам промежуточного и среднего мозга, в том
числе и к красному ядру, где начинается красноядерно—спинномозговой путь
экстрапирамидной системы двигательной регуляции.
Другой важный эфферентный путь идет
от внутреннего отдела бледного шара к передневентральному и вентролатеральному
ядрам таламуса, а оттуда продолжается к двигательным областям коры головного
мозга. Наличие этого пути обусловливает многозвенную петлеобразную связь между
сенсомоторными и двигательными областями коры, которая осуществляется через
полосатое тело и бледный шар к таламусу. Примечательно, что в составе этого
стриопаллидоталамокортикального пути базальные ядра выполняют роль афферентного
звена по отношению к моторным областям коры головного мозга. Многочисленные
связи стриопаллидарной системы с различными отделами мозга свидетельствуют об
ее участии в процессах интеграции, однако до настоящего времени в знании о
функциях базальных ядер остается много невыясненного.
Базальные ядра играют важную роль в регуляции
движений и сенсомоторной координации. Известно, что при повреждении
полосатого тела наблюдается атетоз — медленные червеобразные движения
кистей и пальцев рук.
Дегенерация клеток этой структуры
вызывает также другое заболевание — хорею, выражающуюся в судорожных
подергиваниях мимических мышц и мускулатуры конечностей, которые наблюдаются в
покое и при выполнении произвольных движений. Однако попытки выяснить этиологию
этих явлений в экспериментах на животных не дали результатов. Разрушение
хвостатого ядра у собак и кошек не приводило к возникновению гиперкинезов,
характерных для описанных выше заболеваний.
Локальное электрическое раздражение
некоторых участков полосатого тела вызывает у животных так называемые циркуляторные
двигательные реакции, характеризующиеся поворотом головы и туловища в
сторону, противоположную раздражению. Стимуляция других участков полосатого
тела, напротив, приводит к торможению двигательных реакций, вызванных
различными сенсорными раздражениями.
Наличие определенных расхождений
между данными эксперимента и клиники, по—видимому, свидетельствует о
возникновении системных нарушений механизмов регуляции движений при
патологических процессах в базальных ядрах. Очевидно, эти нарушения связаны с
изменениями функции не только полосатого тела, но и других структур.
В качестве примера можно рассмотреть
возможный патофизиологический механизм возникновения паркинсонизма. Этот
синдром связан с повреждением базальных ядер и характеризуется комплексом таких
симптомов, как гипокинезия — малая подвижность и затруднения при
переходе от покоя к движению; восковидная ригидность, или гипертонус,
независящий от положения суставов и фазы движения; статический тремор
(дрожание), наиболее выраженный в дистальных отделах конечностей.
Все эти симптомы обусловлены
гиперактивностью базальных ядер, которая возникает при повреждении
дофаминергического (по всей вероятности, тормозного) пути, который идет от
черного вещества к полосатому телу. Таким образом, этиология паркинсонизма
обусловлена дисфункцией полосатого тела и структур среднего мозга, которые
функционально объединены в стриопаллидарную систему.
Для выяснения роли базальных ядер в
осуществлении движений успешно используют данные микроэлектродных исследований.
Эксперименты на обезьянах показали наличие корреляции между разрядами нейронов
полосатого тела и медленными, направленными из стороны в сторону червеобразными
движениями лапы. Как правило, разряд нейрона предшествует началу медленного
движения, а при быстрых «баллистических» движениях он отсутствует. Эти факты
позволяют заключить, что нейроны полосатого тела участвуют в генерации
медленных движений, подвергающихся коррекции со стороны сенсорной обратной
связи. Базальные ядра представляют собой один из уровней построенной по
иерархическому принципу системы регуляции движений.
Получая информацию от ассоциативных
зон коры, базальные ядра участвуют в создании программы целенаправленных
движений с учетом доминирующей мотивации. Далее соответствующая информация от
базальных ядер поступает в передний таламус, где она интегрируется с информацией,
приходящей от мозжечка. Из таламических ядер импульсация достигает двигательной
коры, которая отвечает за реализацию программы целенаправленного движения через
посредство нижележащих стволовых и спинальных двигательных центров. Так в общих
чертах можно представить место базальных ядер в целостной системе двигательных
центров мозга.
3.10.1. Морфофункциональная
организация коры больших полушарий
Полушария большого мозга состоят из
белого вещества, покрытого снаружи серым, или корой, толщина которой в
различных отделах больших полушарий колеблется от 1,3 до 4,5мм. Кора
представляет собой филогенетически наиболее молодой и вместе с тем сложный
отдел мозга, предназначенный для обработки сенсорной информации, формирования
двигательных команд и интеграции сложных форм поведения. Бурный рост
неокортекса у высших позвоночных в ограниченном объеме черепа сопровождается
образованием многочисленных складок, увеличивающих общую площадь коры, которая
у человека составляет 2200 см2. На этом пространстве
сконцентрировано 109—1010 нейронов и еще большее
количество глиальных клеток, выполняющих ионорегулирующую и трофическую
функции. Образующие кору нейроны по своей геометрии и функции подразделяются на
несколько групп. Одну группу составляют варьирующие по размеру пирамидные
клетки. Они ориентированы вертикально по отношению к поверхности коры и
имеют тело треугольной формы (рис. 3.30). От тела пирамидной клетки вверх
отходит длинный Т—образно ветвящийся апикальный дендрит, а вниз от основания
нейрона — аксон, который либо покидает кору в составе нисходящих путей, либо
направляется к другим зонам коры. Апикальные и более короткие базальные
дендриты пирамидных клеток густо усеяны мелкими (до 3 мкм) выростами — шипиками,
каждый из которых представляет собой область синаптического контакта.
Другая группа корковых нейронов
представлена более мелкими звездчатыми клетками. Эти клетки имеют
короткие сильно ветвящиеся дендриты и аксоны, формирующие внутрикорковые связи.
Дендриты звездчатых клеток также могут быть снабжены шипиками, которые в
процессе онтогенетического развития у человека появляются только к моменту
рождения.
Наконец, третья группа корковых
нейронов включает в себя веретеновидные клетки, имеющие длинный аксон,
который ориентирован в горизонтальном или вертикальном направлении. В связи с
тем что тела и отростки описанных выше нейронов имеют упорядоченное
расположение, кора построена по экранному принципу и у млекопитающих в типичном
случае гистологически состоит из шести горизонтальных слоев (рис. 3.31).
Самый наружный молекулярный слой
слагается из густого сплетения нервных волокон, лежащих параллельно поверхности
корковых извилин. Основную массу этих волокон составляют ветвящиеся апикальные
дендриты пирамидных клеток нижележащих слоев. Сюда же в наружный слой приходят
афферентные таламокортикальные волокна от неспецифических ядер таламуса,
регулирующих уровень возбудимости корковых нейронов.
Второй слой — наружный зернистый
— состоит из большого количества мелких звездчатых клеток, которые в
вентральной части слоя дополняются малыми пирамидными клетками.
Третий слой — наружный пирамидный
— формируется из пирамидных клеток средней величины. Функционально второй и
третий слои коры объединяют нейроны, отростки которых обеспечивают корково—корковые
ассоциативные связи.
Четвертый слой — внутренний
зернистый — содержит множество звездчатых клеток (клеток—зерен),
обусловливающих его гранулярную структуру. В этом слое преимущественно
оканчиваются афферентные таламо—корковые волокна, идущие от специфических
(проекционных) ядер таламуса.
|
|
|
Рис. 3.30 Структурный микромодуль сенсомоторной коры III—VI
— корковые слои; 1 — интернейроны, 2 — афферентный вход,
контактирующий с интернейроном, 3 —
афферентный вход, образующий терминала на пирамидных нейронах, 4 — возвратная коллатераль аксона,
вступающая в контакт с тормозным интернейроном, 5 — пучок аксонов, выходящий
за пределы колонки, 6 — возвратная
коллатераль, обеспечивающая облегчающие влияния в пределах модуля. 7 —
корково—спинномозговые пирамидные клетки, 8 — корково—красноядерная пирамидная клетка. |
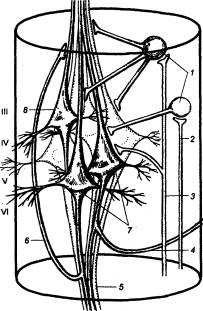
|
|
|
Рис. 3.31 Шестислойное строение коры большого мозга |
Пятый слой — внутренний
пирамидный — образован крупными пирамидными клетками. Наиболее крупные
пирамидные нейроны — гигантские клетки Беца — встречаются в
прецентральной извилине, занятой моторной зоной коры больших полушарий. Аксоны
этих эфферентных корковых нейронов формируют корково—спинномозговой
(пирамидный) и корково—бульбарный пути, участвующие в координации
целенаправленных двигательных актов и позы.
И наконец, шестой слой — мультиформный,
или слой веретеновидных клеток, переходящий непосредственно в белое вещество
больших полушарий. Этот слой содержит тела нейронов, чьи отростки формируют
корково—таламические пути.
Такой шестислойный план строения
характерен для всего неокортекса (рис. 3.32). Однако выраженность отдельных
слоев в различных областях коры не одинакова. Учитывая эту особенность,
немецкий нейроморфолог К. Бродман по гистологическим признакам, в частности по
плотности расположения и форме нейронов, разделил всю кору на 11 областей и 52 цитоархитектонических
поля (рис. 3.33). Позднее были разработаны функциональные принципы классификации
различных зон коры. При этом оказалось, что зоны, выделенные на основании их
функциональных и нейрохимических особенностей, в известной степени
соответствуют цитоархитектоническому разделению коры на поля.
|
|
|
Рис. 3.32 Сильно упрощенная схема нейронов, их сетей, афферентных и
эфферентных связей в различных слоях коры большого мозга А — локализация и форма двух основных типов корковых
нейронов; В —
корково—корковые входы и выходы (ассоциативные а комиссуральные волокна); В —
таламо—корковые (неспецифичные и специфичные) и корково—таламические связи; Г — синаптические входы пирамидной
клетки, аксон которой идет к субталамическим отделам (стволу мозга и спинному
мозгу); Д — обобщенная схема
корковых контуров. |
Так, например, при сравнении наиболее
изученных сенсорных и моторных зон коры оказалось, что в первых наружный
пирамидный слой выражен слабо и доминируют зернистые слои, где оканчиваются
сенсорные афференты (гранулярная кора). И напротив, в моторных зонах
коры зернистые слои развиты плохо (агранулярная кора), а пирамидные слои
превалируют.
Таким образом, функциональная
специализация накладывает определенный отпечаток на структуру сенсорных и
моторных зон коры, и выделение этих областей по различным системам
классификации не случайно.
3.10.2. Проекционные
зоны коры
Использование методов удаления и
раздражения отдельных зон коры позволило идентифицировать ее моторные
(мотосенсорные) области. Важнейшая двигательная область коры локализована в предцентралъной
извилине (поля 4 и б, по Бродману), лежащей кпереди от центральной
(роландовой) борозды, которая отделяет лобную долю от теменной (рис. 3.34). Слабое электрическое
|
|
|
Рис. 3.33 Цитоархитектоническая
карта коры головного мозга человека по Бродману А— верхнелатеральная поверхность; Б — медиальная поверхность. |
раздражение конкретных точек
предцентральной извилины вызывает сокращение определенных групп мышц на
противоположной стороне тела.
В 30—х гг. XX столетия У. Пенфилд, базируясь
на результатах, полученных во время нейрохирургических операций, установил
наличие правильной пространственной проекции соматических мышц различных
отделов тела на двигательную область коры. Обнаружив соматотопическую
организацию двигательной коры, У. Пенфилд отметил, что зоны, управляющие
движениями кистей рук и мимической мускулатурой лица, являются наиболее
обширными и характеризуются самым низким порогом. В дальнейшем на медиальной
поверхности полушария рядом с этой первичной моторной областью была обнаружена
еще одна двигательная область и, таким образом, было показано множественное
представительство периферических образований в нескольких двигательных зонах
коры. В связи с тем, что эти области помимо моторного выхода из коры имеют
самостоятельные сенсорные входы от кожных и мышечных рецепторов, они были
названы первичной и вторичной мотосенсорной корой (МсI и McII).
|
|
|
Рис. 3.34 Упрощенная карта разделения латеральной коры головного мозга на
функциональные поля по Клайсту Цифрами обозначены
цитоархитектонические поля Бродмана (см. рис. 3.33). |
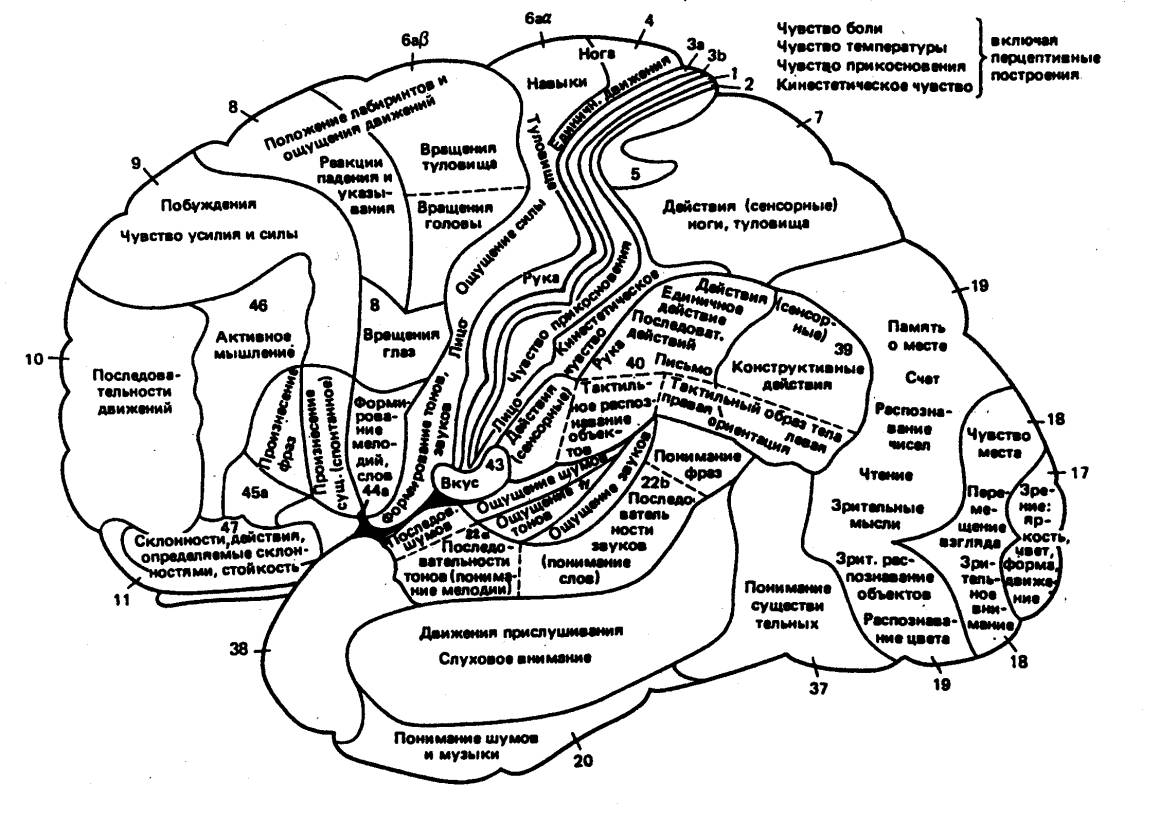
Последующие экспериментальные
исследования показали, что в лежащей позади центральной (роландовой) борозды постцентральной
извилине (поля 1, 2, 3) находится первая соматосенсорная
(сенсомоторная) область, куда через специфические ядра таламуса приходят
афферентные проекции от рецепторов кожи и двигательного аппарата (см. рис.
3.33). Так же как и в двигательной коре, эти проекции имеют соматотопическую
организацию, причем близкие друг от друга точки на поверхности тела имеют
представительство в соседних участках постцентральной извилины. Удаление
определенных зон постцентральной извилины приводит к утрате дискриминационной
чувствительности в тех или иных частях тела. Электрическая стимуляция постцентральной
извилины у больных при нейрохирургических операциях (под местным наркозом)
вызывает чувство онемения или покалывания в определенных местах поверхности
тела.
Кроме первой соматосенсорной области
у хищников и приматов обнаружена вторая соматосенсорная область,
локализованная вентральное в районе латеральной (сильвиевой) борозды,
которая отделяет височную долю от теменной и лобной. Здесь также имеется
соматотопическая проекция поверхности тела, однако менее четкая, чем в первой
соматосенсорной области. Оказалось, что первая и вторая соматосенсорные области
кроме афферентных входов содержат моторные выходы и, следовательно, их
правильнее называть первичной и вторичной сенсомоторными зонами
(См I и См II).
Следовательно,
можно судить о наличии в коре четырех чувствительных областей (См I, См II, МсI и McII), расположенных здесь в порядке убывания их
значимости. Равным образом в коре существует четыре двигательные области (McI, McII, СмI, СмII), причем
ведущая роль в этом ряду принадлежит первичной мотосенсорной коре.
Из числа других проекционных зон
коры следует отметить первичную зрительную область (поле 17),
локализованную в затылочной доле. В проекциях рецепторов сетчатки на
поле 17, так же как и в других сенсорных системах, обнаруживается точный
топографический порядок, причем в каждое полушарие проецируются одноименные
половины сетчаток. Происходящее при этом в каждом полушарии совмещение
зрительных полей обоих глаз лежит в основе бинокулярного зрения. Повреждение
ограниченной области зрительной коры может привести к частичной слепоте,
проявляющейся в нарушении небольшой части поля зрения. Электрическое
раздражение зрительной коры может вызвать зрительные ощущения.
Радом с первичной зрительной
областью расположена вторичная, занимающая поля 18 и 19. Эти поля
имеют отношение к таким функциям, как зрительное внимание и управление
движениями глаз (см. разд. 4.8.4).
В настоящее время хорошо изучена первичная
проекционная зона слухового анализатора, которая локализована в верхнем
крае височной доли (поля 41 и 42). Для этой области, так
же как и для других проекционных зон, описано тонотопическое представительство
спирального (кортиева) органа улитки, состоящее в том, что различные участки
органа проецируются в определенные локусы слуховой коры (см. разд., 4.5.2). Повреждение полей 41 и
42 затрудняет восприятие речи, пространственную локализацию источника
звука и распознавание временных характеристик звука.
В пределах каждой доли коры больших
полушарий (рис. 3.35) рядом с проекционными зонами расположены поля, которые не
связаны с выполнением какой—либо специфической сенсорной или моторной функции.
Такие поля составляют ассоциативную кору, для нейронов которой
свойственно отвечать на раздражение различных модальностей и таким образом
участвовать в интеграции сенсорной информации и в обеспечении связей между
чувствительными и двигательными зонами коры. Эти механизмы являются физиологической
основой высших психических функций.
В состав ассоциативный коры входит
ряд областей теменной, височной и лобной долей. К сожалению, границы этих
областей и корково—корковые связи между ними идентифицированы не полностью, а
большинство физиологических
|
|
|
Рис. 3.35 Четыре основные доли коры головного мозга
(лобная, височная, теменная и затылочная). Вид сбоку 1 — префронтальная ассоциативная кора, 2 — премоторная
кора, 3 — первичная двигательная кора, 4 — центральная борозда, 5 — первичная
соматосенсорная кора, 6 — теменная доля, 7 — вторичная зрительная кора, 8 —
затылочная доля, 9 — первичная зрительная кора, 10— теменно—височно—затылочная
ассоциативная кора, 11 — первичная слуховая кора, 12 — вторичная слуховая
кора, 13 — височная доля, 14 — лимбическая ассоциативная кора, 15 —
латеральная борозда, 16 — лобная доля. |
данных базируется
на симптомах поражения ассоциативных областей у животных и на данных
клинических наблюдений.
Вместе с
тем установлено, что теменные ассоциативные поля, которые в процессе
эволюции надстраиваются над зрительной проекционной зоной, участвуют в оценке
биологически значимой информации и в восприятии пространственных отношений
окружающего мира.
Лобные
доли (поля 9—14),
имеющие обширные двусторонние связи с лимбической системой мозга, контролируют оценку
мотивации поведения и программирование сложных поведенческих актов.
Установлено участие лобных долей в управлении движениями. Так, например,
поражение лобных долей вызывает у больных тенденцию к повторению двигательных
актов без видимого соответствия с внешними обстоятельствами.
Эксперименты
на обезьянах показали, что повреждение лобных долей нарушает запоминание
локализации предметов, причем этот процесс связан с дофаминергическими
структурами, которыми богаты лобные доли.
Очевидно,
лобные доли наряду с теменными участвуют в интеграции сведений о пространстве и
времени.
3.10.3. Колончатая организация зон
коры
Применение современных
микроэлектродных методов для изучения функций корковых нейронов в значительной
мере расширило представления о переработке сенсорной информации в неокортексе.
В 1957 г. американский исследователь В. Маунткасл, анализируя ответы клеток в
соматосенсорной (сенсомоторной) коре кошки на стимулы различных модальностей,
обнаружил следующий интересный факт. При погружении микроэлектрода
перпендикулярно поверхности соматосенсорной коры все встречаемые им клетки
отвечали на раздражитель одной и той же модальности, например на легкое
прикосновение к коже или на движение в суставе. Если же электрод погружали под
углом к поверхности коры, то на его пути попадались нейроны с различной сенсорной
модальностью, чередовавшиеся с определенной периодичностью.
На основании этих экспериментальных
фактов пришли к заключению, что соматосенсорная кора организована в элементарные
функциональные единицы — колонки, ориентированные перпендикулярно
поверхности. Диаметр такой колонки определяется горизонтальным распространением
терминалей афферентного таламокортикального волокна и вертикальной ориентацией
дендритов пирамидных клеток. Колонка является элементарным блоком
сенсомоторной коры, где осуществляется локальная переработка информации от
рецепторов одной модальности. Эта гипотеза колончатой организации неокортекса
получила широкое распространение и дала толчок к дальнейшим исследованиям в
этой области.
Согласно современным представлениям,
каждая функциональная колонка сенсомоторной коры состоит из нескольких
морфологических микромодулей, объединяющих пять—шесть гнездообразно
расположенных нейронов. На рис. 3.30 приводится возможная схема микромодуля
сенсомоторной коры. В его состав входит несколько пирамидных клеток, апикальные
дендриты которых максимально сближены и образуют дендритный пучок; в пределах
этого пучка возможны электротонические связи, которые обеспечивают, по
всей вероятности, синхронную работу всего объединения.
К группе ориентированных по вертикали
пирамидных клеток примыкают звездчатые клетки, с которыми контактируют
приходящие к микромодулю таламокортикальные волокна. Некоторые из звездчатых
клеток, выполняющие тормозную функцию, имеют длинные аксоны, распространяющиеся
в горизонтальном направлении. Аксоны пирамидных клеток образуют возвратные
коллатерали, которые могут обеспечивать и облегчающие влияния в пределах
микромодуля (см. рис. 3.30), и тормозные взаимодействия между микромодулями,
контактируя с тормозными интернейронами. Несколько структурных микромодулей,
объединенных горизонтальным ветвлением терминалей специфических
таламокортикальных афферентов, аксоны терминалей и отростков звездчатых клеток
формируют колонку (или макромодуль), диаметр которой достигает 500—1000
мкм. Каждая колонка характеризуется функциональным единством, которое
проявляется в том, что нейроны колонки отвечают на раздражитель одной
модальности.
В пределах одной колонки нейроны
можно дифференцировать по характеру их импульсных ответов на простые и сложные.
У простых нейронов распределение импульсов в ответе на раздражение
адекватного механорецептора идентично ответу самого рецептора. Сложные
нейроны реагируют на периферические стимулы иначе, чем связанные с ними
рецепторы, и дают максимальный ответ только на определенные формы раздражения,
например на движущиеся стимулы. По всей вероятности, сложные нейроны связаны с
более поздней ступенью анализа тактильного восприятия.
Нейрофизиологические исследования
сенсомоторной коры показали, что обычно возбуждение одного из модулей
сопровождается торможением соседних. Очевидно, этот факт свидетельствует о
том, что процесс торможения препятствует иррадиации возбуждения на смежные
модули и, таким образом, усиливает контрастное различение контуров тактильного
раздражения.
В дальнейшем принцип колончатой
организации был подтвержден при изучении других проекционных зон коры.
Обнаружена ретинотопическая организация зрительной коры, которая, так же как и
сенсомоторная, построена по колончатому принципу. В зрительной коре обезьян наблюдается
регулярное чередование колонок, нейроны которых реагируют на оптическое
раздражение либо только правого, либо только левого глаза. Присутствие
глазодоминантных колонок было подтверждено морфологическими методами, например
с помощью аксонного транспорта меченых аминокислот из глаза в зрительную кору
обезьяны. Кроме этих глазодоминантных колонок в зрительной коре обнаружен
другой набор колонок, отличающихся друг от друга осевой ориентацией
рецептивного поля.
Колончатый принцип организации
нейронов присущ и слуховой коре, где колонки дифференцируются по способности
суммировать информацию, идущую от двух ушей. Наконец, в мотосенсорной коре
также были обнаружены колонки, способные возбуждать или тормозить группу
функционально однородных мотонейронов.
Таким образом, колончатая
организация является общей чертой структуры мотосенсорных и сенсомоторных зон
коры.
Возможно, в первичных проекционных
областях этот принцип организации интимно связан с процессами, создающими в
мозгу отображение различных параметров внешнего раздражителя. Однако вопросы о
том, как используется эта информация за пределами проекционных зон, как из
информации об отдельных качествах внешнего объекта мозг синтезирует цельную
картину реальности, пока остаются открытыми.
По всей вероятности, в мозгу должна
существовать система, которая считывает элементарные процессы в колонках и
объединяет дробную информацию в более сложные сочетания. Реальных данных о
существовании такой системы нейронов нет, однако некоторые гипотезы о возможных
механизмах синтетической деятельности мозга существуют. Одна из них принадлежит
Дж. Эклсу. Согласно этой гипотезе, каждая корковая колонка, или модуль, может
попеременно участвовать в различных сложных системах модулей, аналогичных
динамической констелляции центров по А. А. Ухтомскому.
Формирование такой сложной системы
обусловлено внутрикортикальными связями между отдельными модулями, которые
могут иметь и возбуждающий, и тормозный характер. Активирующие воздействия на
модуль в первую очередь могут поступать через поверхностные горизонтальные
волокна, которые образуют синапсы на апикальных дендритах крупных пирамидных
клеток. Эта активация может усиливаться за счет импульсации, идущей в модуль по
таламокортикальным входам. В то же время через тормозные интернейроны
(например, крупные корзинчатые клетки) активность соседних модулей может быть
подавлена.
Таким образом, баланс возбуждающих и
тормозных процессов в данный момент может выступать в роли координирующего
фактора, который определяет формирование сложных систем модулей. Согласно
современным представлениям, такая система расположенных в разных отделах мозга
и связанных единством функций модулей носит название распределенной.
С точки зрения отмеченных выше
соображений, сегодня старое представление о локализации функций не выдерживает
критики. Даже такой простой поведенческий акт, как захватывание предмета
пальцами, предполагает содружественную работу центров зрительной, моторной коры
и лобной доли, которая участвует в регуляции глазных движений. Вполне естественно,
что управление высшими психическими функциями тем более нельзя считать
компетенцией какого—то одного центра. Реализацию каждой функции осуществляет
распределенная система, состоящая из корковых и подкорковых центров,
соединенных между собой нервными связями.
3.11. ЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В КОРЕ
ГОЛОВНОГО МОЗГА
Электрофизиологические методы нашли
широкое применение для изучения функций коры. У человека и других видов
позвоночных от поверхности головы можно зарегистрировать спонтанные
электрические колебания, обладающие определенной периодичностью. Эти постоянные
колебания отражают электрическую активность коры и обозначаются термином электроэнцефалограмма
(ЭЭГ).
Впервые ЭЭГ у животных была
зарегистрирована в 1925 г. В. Правдич—Неминским, а у людей — немецким
психиатром Г. Бергером в 1929 г.
Для отведения ЭЭГ могут быть
использованы два метода: биполярный и монополярный. При биполярном отведении
оба отводящих электрода расположены на коже головы, т. е. являются активными и
регистрируют разность потенциалов между двумя точками коры. При монополярном
отведении один электрод фиксируют на поверхности головы (активный), а
второй — на мочке уха, это индифферентный электрод. В условиях клиники
расположение электродов при регистрации ЭЭГ стандартизировано и, как правило,
включает отведения от лобных долей, двигательной коры, теменных и затылочных
долей.
|
|
|
Рис.
3.36 Регулярные волны и потенциал (ЭЭГ взрослого человека) А — основные ритмы ЭЭГ: 1 —
α—ритм, 2 — β—ритм, 3 — Θ —ритм, 4 — σ—ритм; Б —
вызванный потенциал в зрительной области коры на световое раздражение
(обозначено стрелкой): 1 — позитивное колебание, 2 — первичное негативное
колебание, 3 — вторичные колебания. |
В острых опытах на животных и при нейрохирургических
операциях на человеке ЭЭГ регистрируют непосредственно от поверхности коры — в
этом случае ее называют электрокортикограммой (ЭКоГ).
Электрокортикограмма имеет более высокую амплитуду и несколько больший
частотный спектр быстрых колебаний по сравнению с ЭЭГ, что обусловлено
исключением дополнительного электрического сопротивления тканей, расположенных
между электродами и поверхностью мозга.
При анализе ЭЭГ с помощью аналоговых
или цифровых устройств учитывают частоту, амплитуду, форму и длительность
слагающих ее электрических колебаний. По этим параметрам в ЭЭГ человека можно
выделить следующие компоненты. У взрослого человека в состоянии покоя и при
отсутствии внешних раздражении в ЭЭГ преобладают регулярные волны, которые
следуют с частотой 8—13 Гц и имеют амплитуду порядка 50 мкВ. Эти волны
обозначаются как α—ритм (рис. 3.36), наиболее выраженный в
затылочных долях коры. α—Ритм возможно регистрировать и от других областей
коры, в частности от теменной доли, где локализована сенсомоторная кора (роландический
ритм).
Переход человека от покоя к
деятельности (восприятие зрительных или акустических сигналов, умственная
работа и т. д.) сопровождается исчезновением α—ритма и возникновением
частых (14—30 Гц) низкоамплитудных (25 мкВ) колебаний β—ритма. Это
явление в физиологической литературе определяется как реакция
десинхронизации ЭЭГ. Синонимами этого термина могут быть реакция
открывания глаз, или реакция активации, характеризующаяся
десинхронизирующим, т. е. тормозящим генерацию α—ритма эффектом.
Если человек из состояния покоя
переходит не к активной деятельности, а, наоборот, ко сну, то в его ЭЭГ
появляются более медленные и высокоамплитудные по сравнению с α—ритмом
волны, в частности Θ—ритм (4—7 Гц) и σ—ритм (0,5—3,5
Гц). Амплитуда этих медленных ритмов варьирует от 100 до 300 мкВ. В норме у
бодрствующих взрослых людей Θ — и σ—ритм не выявляются.
Исключение в данном случае может составлять гиппокампальная кора, где
доминирует Θ —ритм при активном состоянии.
Таким образом, анализ частотного спектра
ЭЭГ позволяет довольно четко судить о функциональном состоянии коры и не
случайно электроэнцефалографию широко используют в клинике. Травмы и
патологические процессы в мозгу иногда вызывают специфические изменения ЭЭГ, по
которым можно установить локализацию болезненного очага. Так, например, для
больных эпилепсией характерно наличие в ЭЭГ судорожных пиковых разрядов и
специфических пароксизмальных волн, возникающих в патологической зоне.
Вопрос о генезисе волны ЭЭГ является
довольно сложным; вместе с тем можно утверждать, что волны ЭЭГ являются
результатом алгебраической суммации постсинаптических потенциалов корковых
нейронов. Наиболее эффективная суммация происходит при синхронном возбуждении
многих клеток, которое проявляется при ограничении сенсорного притока. Приход
сенсорной импульсации в кору (например, при открывании глаз) расстраивает
синхронизацию и приводит к смене α—ритма на β—ритм, или к реакции
десинхронизации ЭЭГ.
Причиной возникновения синхронных
постсинаптических изменений в корковых нейронах могут быть циклические
таламокортикальные взаимодействия в которых таламические нейроны играют роль
своеобразных ритмоводителей или пейсмекеров. В пользу этой точки зрения
свидетельствуют следующие экспериментальные факты. В таламических центрах обнаружена
ритмическая активность, частота которой совпадает с α—ритмом. После
экспериментального нарушения таламокортикальных связей α—ритм в коре
исчезает, а в таламических структурах сохраняется.
Одним из возможных механизмов
ритмической активации таламических нейронов считают возвратное самоторможение,
создающее периодические колебания возбудимости. Кроме того, на генерацию
таламического ритма влияют импульсы, поступающие из ретикулярной формации
ствола. Частота этих импульсов зависит от афферентного притока в
неспецифическую систему ретикулярной формации, которая может и стимулировать, и
тормозить ритмическую активность таламических центров.
3.11.2. Метод вызванных потенциалов
При исследовании функций коры
успешно используют метод вызванных потенциалов. Вызванные потенциалы
представляют собой регистрируемые с поверхности коры электрические реакции в
ответ на раздражение рецепторов, периферических нервов, таламических ядер и
других образований, связанных с проведением сенсорных сигналов.
Вызванные потенциалы принято
разделять на две группы: первичные ответы (ПО) и вторичные ответы (ВО). Первичные
ответы, представляют собой двухфазные, позитивно—негативные (см. рис. 3.36)
колебания, которые регистрируются в проекционных зонах коры при раздражении
специфических афферентов; ПО характеризуются коротким скрытым периодом,
стойкостью к наркозу и способностью воспроизводить довольно высокий ритм (до
120 имп/с) афферентных сигналов. Первичные ответы являются локальными
процессами и регистрируются только в пределах проекционной зоны. Это свойство
ПО было использовано при исследовании топической организации различных
проекционных зон и при идентификации связей между различными структурами мозга.
Первичные ответы возникают за счет
суммации постсинаптических потенциалов нейронов, расположенных в изучаемом
участке коры. По—видимому, начальная позитивная фаза ПО обусловлена суммацией
синаптических потенциалов тел и базальных дендритов пирамидных клеток.
Негативная фаза связана с активацией апикальных дендритов этих же нейронов в
поверхностных слоях коры.
Позитивная полярность первой фазы ПО
определяется пространственным расположением электрода по отношению к источнику
напряжения и конфигурацией электрического поля вокруг возбужденного нейрона в
коре, являющейся объемным проводником. В результате регистрируемое
поверхностным электродом позитивное колебание ПО отражает деполяризацию тел
пирамидных клеток, локализованных в более глубоких слоях коры. При отведении
негативной фазы ПО электрод максимально приближен к источнику напряжения
(апикальным дендритам), и это исключает изменение полярности, обусловленное
дипольным моментом. Справедливость этой точки зрения подтверждается тем, что
при погружении регистрирующего электрода в кору полярность первой позитивной
фазы ПО изменяется на отрицательную по мере приближения к телам пирамидных
клеток, т. е. к источнику напряжения.
Вторичные ответы (ВО), которые также включают
ассоциативные ответы как самостоятельную категорию, в отличие от ПО довольно
изменчивы по форме, обладают большей чувствительностью к наркозу, большим
скрытым периодом и исчезают при частоте раздражения более 5 имп/с. Большинство
из них не обладает узкой локализацией и регистрируется в ассоциативных корковых
зонах, примыкающих к проекционным. Вторичные ответы, возникающие при
раздражении неспецифических структур мозга (ретикулярная формация мозга,
неспецифические ядра таламуса), имеют генерализованный характер и могут быть
зарегистрированы в коре повсеместно. Функционально ВО отражают процессы,
связанные с передачей возбуждения от первичных проекционных зон к ассоциативным
и с рабочей настройкой корковых нейронов путем изменения их возбудимости.
Метод регистрации вызванных
потенциалов может быть использован при анализе процессов, связанных с
формированием условного рефлекса. Одним из показателей этих процессов является
описанная Г. Уолтером волна ожидания, или условное негативное
отклонение. Если испытуемому дается инструкция нажимать кнопку на второй из
двух последовательных сигналов, то после ряда сочетаний первый раздражитель
(звуковой щелчок) начинает вызывать волну ожидания, которая продолжается до
подачи второго раздражителя (вспышка света).
Волну ожидания регистрируют от
поверхности головы в зоне лобных и теменных долей, она представляет собой
медленное негативное колебание с амплитудой порядка 40 мкВ. Волна ожидания
предшествует движению, которое должен произвести испытуемый, и, по—видимому,
является электрофизиологическим отражением процессов подготовки к
целенаправленному движению. Однако детально связь волны ожидания с психическими
процессами человека пока не расшифрована.
Использование микроэлектродных
методов исследования позволило детально изучить биофизические свойства
одиночных пирамидных клеток, которые отличаются сравнительно большими размерами
и могут быть идентифицированы. Оказалось, что основные электрофизиологические
характеристики пирамидных клеток сходны с таковыми у мотонейронов. Потенциал
покоя колеблется в пределах от 60 до 80 мВ; потенциал действия достигает 60—100
мВ и имеет длительность 0,5—2 мс. Он возникает в низкопороговой зоне клетки
(аксонном холмике) и оттуда распространяется по аксону и антидромно, попадая в
тело и дендриты пирамидных нейронов. Постсинаптические потенциалы пирамидных
клеток длительнее, чем у мотонейронов. Особенно это касается ТПСП,
продолжающихся 70—150 мс и дольше.
Интегрирующая деятельность
пирамидных нейронов, так же как и других центральных нейронов, обусловлена
суммацией синаптических потенциалов на клеточной мембране.
3.12. ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ
Основной вопрос при рассмотрении
эволюции мозговых систем поведения целостного организма состоит в том, может ли
изучение ныне живущих животных дать достоверные сведения о тех существах,
которые жили десятки миллионов лет назад. Обращаясь к филогенетическому
древу, можно видеть, что по мере развития основного ствола млекопитающих от
него отходит ветвь современных однопроходных, позже сумчатых, стволом же
плацентарных млекопитающих являются насекомоядные, от которых произошли
приматы, хищные и грызуны. Поэтому иногда встречающееся сравнение крыс, кошек и
обезьян, якобы представляющих собой филогенетический ряд, неправомерно с точки
зрения истории их развития. Наоборот, большая часть отрядов современных
млекопитающих — результат параллельной эволюции, а не последовательной, к
которой можно отнести однопроходных, сумчатых и насекомоядных.
Особенно важна специфика мозговой
организации насекомоядных, которых рассматривают как потомков общих предков,
давших начало высшим плацентарным. Есть основание считать, что план организации
мозга насекомоядных является предшественником для его дальнейшего
усовершенствования в параллельных независимых друг от друга рядах — грызунов,
хищных и приматов. Филогенетический подход не исключает и другого, адаптационного
подхода, который основан на изучении ныне живущих форм, обладающих разной
степенью адаптации (специализации) в смысле развития мозга и сенсорных систем.
Представители такого сравнительного ряда не обязательно должны иметь общую
линию эволюции, и выводы, сделанные в результате такого сравнения, будут
относиться только к общим принципам адаптации и выживания.
Филогенетический и адаптационный
подходы не исключают, а взаимно дополняют друг друга. Поэтому выделяют пути
филогенетической и адаптивной эволюции.
3.12.1. Происхождение новой коры
У предков млекопитающих, как и у
современных рептилий, кора больших полушарий обладала очень невысоким уровнем
дифференциации. Значительному росту неокортекса млекопитающих
способствует прогресс старой (археокортекс) и древней (палеокортекс) коры.
Некоторые связывают кортикогенез с древней корой, обонятельным мозгом, считая
обонятельную афферентацию ведущей в процессах кортикогенеза.
Другие считают, что неокортекс
произошел в результате дифференциации структур старой, гиппокампальной, коры.
Поэтому формирование новой коры уже у низших млекопитающих связывают не столько
с обонятельной сигнализацией, сколько с представительством всей совокупности
сенсорных систем.
А. А. Заварзин считал, что развитие
новой коры, т. е. слоистых, экранных структур, вызвано у млекопитающих
переключением зрительного пути на кору большого мозга, а поэтому и развитие
новой коры он связывал со зрительной системой. Не менее важная роль как фактора
прогрессивного кортикогенеза принадлежит двигательно— проприоцептивной системе.
Считают вероятным, что зрительная и соматическая афферентные проекции
определили формирование новой коры. Ведущим рассматривают процесс вступления
афферентных волокон из таламуса (зрительного бугра) в древнюю кору на границе
со старой корой и превращение соответствующего участка коры в проекционный
район определенного вида чувствительности. В зарождающуюся кору первыми по
такому пути стали поступать импульсы соматической чувствительности, а затем
зрительные и другие импульсы.
Экранная структура новой коры
явилась субстратом для проекции и объединения деятельности многих сенсорных
систем. Одновременно развивался и собственный эффекторный аппарат коры — пирамидные
пути. При сопоставлении низших млекопитающих с субприматами и приматами
видно, что диапазон адаптивных возможностей животного тесно коррелирует с
уровнем развития не столько самих специфических сенсорных аппаратов, сколько
мозговых систем, обеспечивающих преимущественно объединение, интеграцию всех
сенсорных влияний. Применительно к млекопитающим это своеобразные структуры
таламуса и коры, не принадлежащие какой—либо сенсорной системе, но в то же
время получающие импульсацию от нескольких сенсорных систем.
На стволовом уровне мозга сложные
интегративные функции приписывают ретикулярной формации —
филогенетически древней неспецифической структуре, из которой формируются
специфические ядра ствола. Аналогичные изменения происходят и с неспецифической
системой таламуса и общей неспецифической корковой пластинкой у пресмыкающихся.
Наряду с формированием специфических реле — ядер таламуса — многие структуры
продолжают оставаться неспецифическими — они получают множество разномодальных
входов и не имеют локальной проекции на ограниченные корковые районы. Из такой
неспецифической системы таламуса начинает выделяться ряд образований, которые
получают импульсацию от ядер разных афферентных систем и проецируют ее в
корковые структуры, располагающиеся между специфическими сенсорными . зонами.
Именно этим системам, которые
именуются ассоциативными, и принадлежит наиболее важная роль в
организации интегративной функции мозга.
3.12.2. Организация новой коры у
низших млекопитающих
Сведения о функциональной
организации коры больших полушарий у однопроходных крайне ограничены.
Электрофизиологическое обследование новой коры у ехидны показало, что основные
сенсорные проекции располагаются в затылочной доле коры позади так называемой
α—борозды. Спереди от нее находится агранулярная кора, а сзади — гранулярная.
Соматосенсорное представительство перекрывается моторным. Не обнаружено зон
перекрытия типа ассоциативных. В верхней части электровозбудимой зоны берет
начало пирамидный тракт, который прослеживается до 24—го спинного сегмента.
Мозолистое тело отсутствует.
Мозг утконосов лишен борозд и
извилин и по плану функциональной организации напоминает мозг ехидны. Моторные
и сенсорные проекции перекрываются не на всем протяжении, тогда как зрительные
и слуховые проекции в затылочном полюсе коры перекрываются между собой и
частично с соматической проекцией. Такая организация неокортекса утконоса,
приближающаяся к корковой пластинке рептилий, позволяет его рассматривать как
еще более примитивный в сравнении с ехиднами.
Следовательно, мозг однопроходных
сохраняет еще многие черты мозга рептилий и в то же время отличается от последних
общим планом строения, характерным для млекопитающих.
Значительно более изучены сумчатые,
общая схема строения таламуса которых в основном соответствует таковой у высших
млекопитающих. Для ядер таламуса обнаружено перекрытие слуховой и соматической
систем. У сумчатых нет мозолистого тела. В отношении общего плана
таламокортикальных проекций сумчатые не представляют исключения из остальных
млекопитающих. Кора мозга также построена по классическому шестислойному типу.
Корковые представительства соматомоторных и соматосенсорных систем почти
полностью перекрываются. Существуют перекрытия и других сенсорных систем, хотя
и не столь выраженные, как у однопроходных.
Пирамидный путь начинается от коры
позади орбитальной борозды и заканчивается на интернейронах спинного мозга на
уровне 7—10—го грудных сегментов. Обнаружены кроме этого другие корковые
нисходящие тракты к ядрам моста и продолговатого мозга, которые можно
рассматривать как предпосылки для дальнейшего развития у высших млекопитающих
корково—луковичного и корково—мостового нисходящих путей.
У сумчатых не обнаружено участка
коры, которая по своим свойствам хотя бы отдаленно напоминала ассоциативную
кору высших млекопитающих.
Плацентарные млекопитающие, низшими из которых являются
насекомоядные, характеризуются близостью общей конструкции мозга к грызунам и
хищным. Хотя насекомоядные характеризуются более развитым неокортексом,
чем сумчатые, мозолистое тело мало чем отличается у представителей обеих групп
животных. Зоны коры с диффузно распределенными по слоям комиссуральными
волокнами являются остатками старой неспециализированной коры, столь
характерной для рептилий. У ежей слуховое и зрительное таламические ядра еще
полностью не дифференцированы и не имеют четкой модальной специфичности. В коре
больших полушарий описан поясок вторичного зрительного поля, расположенный
между первичным зрительным и слуховым полями. На него проецируется заднее
латеральное ядро таламуса, являющееся предшественником подушки зрительного
бугра.
Появление вторичных корковых сенсорных
зон — прогрессивный фактор эволюции. Помимо проекционных зон в коре мозга ежей
обнаружена полисенсорная ассоциативная зона, которая охватывает
архитектонические поля 5 и 7 теменной доли. Эта зона непосредственно
перекрывается со вторичным зрительным и соматическим полями (V—II и S—II), на нее проецируются ассоциативные ядра таламуса
(заднее латеральное и медиодорсальное). По нейронному строению ассоциативная
зона сложнее, чем другие районы коры: это в основном касается высокодифференцированных
клеток верхних слоев, развитой дендритной системы нейронов, пространственного
распределения различных афферентных входов по поперечнику коры (хотя по
электрофизиологическим данным мультисенсорные нейроны располагаются диффузно и не
обнаруживают преобладания того или иного сенсорного входа). Последние были
равноценными по степени активации ассоциативной коры. Пирамидный тракт у
насекомоядных берет начало от гигантских пирамидных клеток (Беца) в передней
теменной и островковой областях коры. Фронтальная зона коры еще не посылает
волокон в пирамидный путь.
Следовательно, у насекомоядных
гигантопирамидное теменное поле является одновременно предшественником
ассоциативных систем мозга и источником волокон пирамидного пути, т. е.
структурой, где осуществляются процессы сенсорной интеграции. Разрушение этой
области неокортекса лишает животное наиболее тонких актов сенсорного контроля
произвольных движений в составе цепного двигательного условного рефлекса.
В параллельных рядах, которыми шло развитие
современных млекопитающих, хотя и сохранился общий план конструкции мозга, но
его таламокортикальные системы претерпели существенные морфофункциональные
перестройки. Высокого развития достигли корковые механизмы деятельности
сенсорных систем с тенденцией возрастания специфических проекций и
ассоциативных корковых полей со свойствами мультисенсорного конвергирования.
Эти отделы мозга как зоны перекрытия корковых сенсорных проекций обособились; с
ними связывают реализацию наиболее сложных форм межсенсорной интеграции.
3.12.3. Организация новой коры у
высших млекопитающих
В отношении функциональной
организации коры грызуны еще мало отличаются от насекомоядных. На
наружной поверхности коры не обнаружено ассоциативных зон, проекция афферентных
систем достаточно диффузна с большими зонами перекрытия, в них преобладает (как
и у ежей) неспецифический тип мультисенсорной конвергенции с равноценностью
сенсорных входов. Все это коррелирует с низким уровнем интегративной
деятельности мозга грызунов, у которых преобладают автоматизированные формы
поведения.
У хищных выделяют два
самостоятельных ассоциативных поля — теменное и лобное, каждое из которых
характеризуется конвергенцией различных сенсорных входов и наличием выходных
волокон в составе пирамидного пути. У кошек и собак таковыми свойствами
обладают теменная и заднелобная области коры (гигантопирамидные поля), а
переднелобная кора как зона широкой конвергенции сенсорных влияний формирует
самостоятельные нисходящие тракты к ядрам моста и продолговатого мозга.
Дальнейший прогресс в конструкции
больших полушарий отмечен у приматов, характеризующихся наибольшим
развитием таламо—теменной и таламо—лобной ассоциативных систем. Эти
ассоциативные корковые поля достигают особенного развития у человека и занимают
более половины всей поверхности неокортекса.
Кроме того, у приматов впервые в
эволюции больших полушарий появляются длинные пучки волокон, соединяющих между
собой все ассоциативные поля коры в единую интегративную систему мозга.
Территория специфических проекционных
зон коры по сравнению с хищными значительно сокращается. У приматов также
территориально обособляются корковые зоны, дающие начало пирамидному пути. При
этом все три ассоциативные зоны неокортекса оснащаются экстрапирамидными
эфферентными путями, посылающими свои волокна к ядрам моста, среднего и
продолговатого мозга и оттуда к мозжечку. Таким путем кора больших полушарий
подчиняет себе деятельность практически всех нижележащих отделов головного
мозга.
Таким образом, прогрессивное
развитие ассоциативных систем мозга, коррелирующее с уровнем совершенства
аналитико—синтетической деятельности, может рассматриваться в качестве
показателя филогенетического статуса вида и уровня его адаптивных возможностей.
Лобная кора — не только коллектор
множества эфферентных влияний, конвергирующих к ней из разнообразных мозговых
источников, но это и мощный аппарат регулирования и управления структурами
мозга. Здесь расположены основные пути саморегуляции внутримозговых систем',
мезэнцефалическая ретикулярная формация г— основное звено управления
функциональным состоянием мозга, ассоциативный таламус — главный докорковый
уровень зиготы до начала родов) периоде развития. Дифференциация нейронов
начинается с глубоких слоев коры. К окончанию миграции нейробласты в глубине корковой
пластинки формируют V—VI слои коры, а те нейробласты, которые мигрировали
последними, — II слой.
Морфологическое созревание нейронов
выражается в появлении и накоплении базофильного вещества (субстанции, или
зернистости, Ниссля), развитии отростков и их ветвлений, росте ядра и тела
клетки, миелинизации аксонов и образовании синапсов. Уже у новорожденных клетки
снабжены отростками, рост и развитие которых продолжается в течение первых лет
жизни.
Одним из критериев созревания коры
является прогрессивное уменьшение плотности клеток. Таламические же
афференты, подрастая к коре, способствуют дифференциации слоев коры и
стимулируют развитие нейронов. Постепенно в течение раннего постнатального
онтогенеза максимум плотности афферентных таламических волокон сдвигается от
крайних слоев к срединным.
Таким образом, все нейроны коры
формируются благодаря митозу, однако это формирование происходит не в одно и то
же время. Нейроны закладываются и мигрируют из глубины коры по направлению к
поверхности (за исключением I слоя, в котором преобладают дендритные ветвления
глубоко расположенных клеток). Мигрирующие клетки могут образовывать
синаптические контакты с клетками, мимо которых они проходят к местам
окончательного расположения. Звездчатые клетки к моменту рождения еще
продолжают мигрировать, а пирамидные клетки V—VI слоев уже к этому времени
занимают свои окончательные позиции.
3.12.5. Развитие корковых межнейронных связей
В основном межнейронные синапсы
проходят путь своего развития в постнатальном онтогенезе. У новорожденных
животных незрелость ультраструктуры проявляется в небольшой протяженности
активных зон синаптических мембран, малом количестве синаптических пузырьков,
диффузно распределенных в пресинаптических терминалях. К концу 1—й недели
повышается электронная плотность в местах контакта мембран, увеличивается
количество везикул, их концентрация в области активных зон, в синаптической
щели появляется субсинаптическая сеть. И только к 2—недельному возрасту
отмечается сходство со строением синапсов взрослых. Среди аксодендритических
контактов раньше образуются синапсы с круглыми, а позднее — с удлиненными
везикулами, что предположительно связывают с разными сроками созревания
возбуждающих и тормозных синапсов.
В постнатальном онтогенезе особенно
важен этап прозревания животного. С открыванием глаз резко увеличивается
электронная плотность синаптических мембран, пузырьки группируются в местах их
активного контакта. Количество синапсов существенно зависит от степени зрелости
той или иной области коры: если у слепых новорожденных крысят в зрительной коре
зрелые синапсы отсутствуют, то в сенсомоторной коре они обнаруживаются в
большом количестве. Число синапсов зависит от формирования системы дендритов и
коллатералей аксонов, т. е. от функциональной нагрузки коры мозга.
Раньше
всего синапсы появляются на основном стволе дендрита, затем на больших ветвях
дендритов и позднее на их более тонких ветвях. Позднее всех созревают
аксошипиковые синапсы. Именно в этот период отмечено интенсивное образование аксосоматических
синапсов. Предполагают, что рано созревающие контакты являются стабильными, а
позднее формирующиеся — более изменчивые и динамичные. Последние в большей мере
ответственны за пластические свойства коры головного мозга.
Различные зоны коры больших
полушарий формируются в разные сроки. Так, у кошек общая нейронная и
синаптическая организация теменной коры обнаруживает признаки зрелости раньше,
чем сенсомоторная кора. Первой в сенсомоторной коре у кошек созревает наиболее
биологически важная рефлекторная система, обеспечивающая
координированное управление движениями передних конечностей и лицевой
мускулатуры при выполнении реакции сосания. Это соответствует правилу
системогенеза, согласно которому созревание различных рефлекторных актов
происходит гетерохронно в зависимости от их биологической значимости. Далее
наблюдается бурный рост внутрикортикальных, кортикоталамических связей и связей
коры с другими отделами мозга.
Каждый нейронный модуль
сенсомоторной коры становится потенциально полифункциональной единицей,
способной принимать сигнализацию от различных отделов мозга и от разных
рецепторных зон. Наконец, формируется параллельно, но с некоторым запаздыванием
внутрикорковая тормозная система, Она способствует не только специализации
корковых модулей в смысле ограничения притока к ним афферентной импульсации, но
и устанавливает между ними сложные координационные отношения. Образуется
рабочая динамическая система модулей, обеспечивающая корковое управление
целостным поведенческим актом.
3.13. НАСЛЕДСТВЕННО ЗАКРЕПЛЕННЫЕ
ФОРМЫ ПОВЕДЕНИЯ
Различные формы поведения принято
разделять на врожденные и приобретенные в процессе
индивидуального развития. И те, и другие направлены на сохранение индивидуума и
вида. В процессе филогенеза сформировались механизмы, обеспечивающие
функциональное единство отдельных систем организма и поддерживающие его
взаимодействие с внешней средой. На определенном этапе эволюции возникла
рефлекторная деятельность — ответы на воздействия окружающей или внутренней
среды, опосредованные через центральную нервную систему. Наследственно
закрепленные рефлексы лежат в основе приспособительных поведенческих актов,
проявляющихся без предварительного научения. Они видоспецифичны, т. е.
неизменны у всех представителей данного вида в ответ на действие
соответствующих им раздражителей внешней среды при определенном состоянии
организма.
Организм приспособлен к условиям
окружающей среды на всем протяжении онтогенеза, включая и антенатальный период
развития. В соответствии с развертыванием генетической программы отдельным
этапам онтогенеза свойственны характерные комплексы движений. У плодов
млекопитающих по мере созревания ЦНС вначале возникают обобщенные, а затем
специализированные рефлекторные реакции. К моменту рождения созревают комплексы
центральных и периферических нервных образований и связанных с ними эффекторных
аппаратов, обеспечивающих существование вне организма матери. В постнатальной
жизни последовательно проявляются вначале менее сложные, далее все более
сложные специализированные поведенческие акты. Наряду с ними совершенствуются и
вегетативные реакции, связанные с поддержанием гомеостаза.
В период новорожденности происходит
перестройка рефлекторной деятельности, некоторые формы поведения устраняются и
заменяются новыми. В пренатальном и в постнатальном периодах развития
проявляются элементы и комплексы поведения разной сложности, направленные на
удовлетворение различных потребностей организма. Задача физиологии — выявить
механизмы врожденного поведения и его взаимодействий с процессами научения.
Исследования, проводимые в этом направлении, традиционны для отечественной
физиологии и восходят к представлениям И. М. Сеченова о рефлекторной основе
психической деятельности. В русле рефлекторной теории И. П. Павлов
рассматривал врожденное поведение как совокупность сложнейших безусловных
рефлексов (инстинктов).
С несколько иных позиций
разрабатывают проблему наследственно закрепленного поведения зоопсихологи, а в
последние 50 лет и этологи, которых интересуют преимущественно общебиологические
основы инстинктов.
Однако в настоящее время наблюдается
сглаживание противоречий между физиологическим, зоопсихологическим и
этологическим подходами к изучению инстинктов и попытка создать синтетическую
теорию, охватывающую различные стороны врожденного поведения.
Прежде чем перейти к изложению
современных представлений о наследственно закрепленных формах поведения,
целесообразно остановиться на их исторических
предпосылках.
С давних времен было замечено, что в
естественной среде животные хорошо приспособлены к условиям существования и
обладают специфическим набором поведенческих актов, присущих данному виду и
проявляющихся с самого рождения. Научный анализ инстинктов был впервые
предпринят в XVIII в. французским врачом, философом—материалистом Ж. Ламетри,
который рассматривал их как действия, независимые от разума и опыта. Это
определение основывалось на сравнительном анализе приспособительных актов
млекопитающих, птиц, рыб и насекомых.
Ч. Дарвин, определивший инстинкт как
отражение наследственных свойств мозга, проанализировал эволюцию
инстинктивной деятельности. Он показал, что формы адаптивного поведения
складываются в процессе эволюции данного вида, и привел важные доказательства
роли отбора в появлении приспособительных видоспецифических врожденных стереотипов
поведения на примерах строительного инстинкта пчелы, гнездового паразитизма
кукушки, «рабовладения» у некоторых видов муравьев и на основе сопоставления
поведения диких и домашних животных.
В России идеи Ч. Дарвина получили
широкое развитие. В. А. Вагнер на основании полевых наблюдений и экспериментов
не только показал роль среды и естественного отбора в возникновении
инстинктивных форм поведения, но и выдвинул положение об их пластичности,
определяемой внешними условиями. Соотношение врожденного и индивидуально
приобретенного в процессе воспитания, т. е. роль опыта в изменении
инстинктивной деятельности, изучали И. П. Павлов, его сотрудники и
последователи. Л. А. Орбели проанализировал различия в пластичности врожденного
поведения зрело— и незрелорождающихся животных. А. Д. Слоним на основании
исследования условных и натуральных условных рефлексов в связи со средой
обитания и образом жизни выдвинул положение о сложнорефлекторных формах
поведения. Л. В. Крушинский ввел понятие унитарные реакции поведения, в
которых интегрированы условные и безусловные рефлексы. Сравнительная
архитектоника (от греч. architektonike — построение, структура) врожденных
и приобретенных поведенческих актов была проанализирована в исследованиях В. М.
Бехтерева, И, С. Бериташвили, Н. А. Рожанского и многих других.
Эволюционные аспекты взаимоотношения
инстинктивного и индивидуально приобретенного поведения разработаны А. Н.
Северцовым, который оценивал инстинкты как
устойчивые приспособления, столь же важные для существования вида, как и
морфологические признаки. Если условные рефлексы не наследуются, а определяются
врожденным уровнем организации мозга, то в инстинктивном поведении
наследственно закреплены фиксированные программы действий и способность их к
определенной модификации в результате индивидуального опыта.
Одним из основных направлений в
изучении врожденного поведения является этология, изучающая поведение
животных, начиная с беспозвоночных и кончая антропоидами и даже человеком в адекватной
для них среде. Описание и сравнительный анализ характерных для разных видов
поведенческих актов (этограмм) проводится в русле этого направления с
использованием хронометража, фото— и киносъемки, видеомагнитофонных записей
поведения. Основоположник этологии зоолог К. Лоренц, а также Н. Тинберген и К.
фон Фриш за исследования в области поведения животных в 1973 г. были удостоены
Нобелевской премии.
Если в наше время сглаживаются
различия в подходе к изучению поведения животных у биологов разных специальностей
— физиологов, психологов, генетиков, эволюционистов — и делаются попытки
создания синтетической теории, то в начале XX века противоречия между ними были
еще весьма значительны.
И. П. Павлов противопоставил
рассмотрению поведения животных в антропоморфическом аспекте учение о высшей
нервной деятельности, положив в его основу понятие об условном рефлексе.
Это учение определило успешно развивающийся и в наше время физиологический
подход к изучению центральных механизмов поведения.
В начале нашего века сформировалось
новое течение в психологии — бихевиоризм (от англ. behavior — поведение), представители
которого также отвергали интроспекцию в изучении поведения, стремясь к
максимальной его объективизации, однако мало интересовались его центральными
механизмами. Тем не менее американский психолог Дж. Уотсон, основатель
бихевиоризма, указывал на то, что именно представление об условном рефлексе
дало побудительный толчок к созданию его теории. Он сформулировал несколько
важных принципов объективного изучения поведенческих актов, описывая поведение
без попытки объяснить его внутреннюю природу.
Согласно схеме бихевиористов,
деятельность животного организма строится по декартовскому принципу
«стимул—реакция». В дальнейшем бихевиористы ввели в эту схему «промежуточные
переменные» — силу навыка, величину потребности, утомление и ряд других
побудительных факторов. Бихевиористы разработали ряд объективных методик
изучения поведения животных, в частности проблемные клетки, лабиринты, камеры
для исследования инструментальных реакций и др.
В наше время большинство физиологов,
изучающих поведение, используют весь арсенал методических подходов, позволяющих
вскрывать его механизмы. При этом основной акцент делается на исследовании процессов
научения. Между тем их структурную организацию можно понять лишь на
основании достаточных сведений о наследственно закрепленной деятельности. С
этой точки зрения необходимо исходить из предложенного И. П. Павловым
разделения нервной деятельности на низшую — безусловные рефлексы — и высшую
— условные рефлексы.
Безусловные рефлексы были выделены в
специальную категорию для обозначения видовых реакций организма на внутренние и
внешние раздражители, осуществляющихся на основе врожденных нервных связей, т.
е. отражающих филогенетический опыт приспособления к условиям существования.
Безусловные рефлексы относительно постоянны, стереотипно проявляются в ответ на
адекватное раздражение определенного рецептивного поля и служат основой для
формирования многочисленных условных рефлексов, связанных с индивидуальным
опытом. Безусловные рефлексы , обеспечивают координированную деятельность,
направленную на поддержание постоянства многих параметров внутренней среды,
взаимодействие организма с внешней средой, согласованную деятельность
соматических, висцеральных и вегетативных реакций.
Однако оптимальное приспособление
к/меняющимся состояниям внешней и внутренней сред организма достигается с
помощью условных рефлексов, благодаря которым индифферентные для определенной
деятельности раздражители приобретают качество биологически значимых сигналов.
Предложено несколько классификаций
безусловных рефлексов в соответствии с характером вызывающих их
раздражителей, их биологической ролью, уровнями управления (связь с
определенными отделами ЦНС), порядком следования в конкретном приспособительном
акте. Авторы этих классификаций отразили в них научные интересы и
методологические установки. И. П. Павлов описал пищевые, оборонительные,
ориентировочные, родительские и детские реакции, подразделяемые на более
дробные рефлексы. Так, пищевые рефлексы, связанные с деятельностью пищевого
центра, включают поиск, добычу, захват, вкусовое опробование пищи, секрецию
слюны и пищеварительных соков в желудочно—кишечном тракте, его моторную
деятельность.
В трудах И. П. Павлова встречаются
также упоминания следующих безусловных рефлексов: пищевые (положительный и
отрицательный), ориентировочный, коллекционирования, цели, осторожности,
свободы, исследовательский, самосохранительные (положительный и отрицательный),
агрессивный, сторожевой, подчинения, половые (мужской и женский), игровой,
родительский, норо—гнездный, миграционный, социальный, питьевой.
Н. А. Рожанский выделил 24 рефлекса,
входящие в следующие шесть групп:
общей активности, обменные,
междуживотных отношений, продолжения вида и размножения, экологические и
неповеденческие рефлексы подкорково—стволовых частей головного мозга. Эта
классификация почти не затрагивает вегетативную сферу регуляции, играющую
большую роль в реализации поведенческих актов.
Более широкой является
классификация, основанная на исследовании адаптивных сторон
безусловнорефлекторной деятельности. Представитель эколого—физиологического
направления А. Д. Слоним предложил разделить безусловные рефлексы на три
группы реакций, связанных с поддержанием постоянства внутренней среды,
изменениями во внешней среде и сохранением вида.
Приведенные классификации
предусматривают не только описание поведения, но и выяснение лежащих в его
основе физиологических механизмов. Последнее в меньшей степени интересует
этологов, также исследующих поведение в адекватной для животного окружающей
среде.
Для П. В. Симонова классифицирующим
принципом группировки сложнейших безусловных рефлексов явились идеи В. И.
Вернадского и А. А. Ухтомского об освоении живыми существами разных уровней
организации в гео—, био—, а для человека также в социо— и ноосфере
(интеллектуальное освоение мира). П. В. Симонов выделил следующие безусловные
рефлексы: витальные, ролевые (зоосоциальные) и саморазвития. К витальным
безусловным рефлексам относятся пищевой, питьевой, регуляции сна,
оборонительные (включая рефлекс «биологической осторожности»), рефлекс экономии
сил и многие другие. Они не требуют участия другой особи, а невозможность их
реализации ведет к физической гибели. Ролевые (зоосоциальные)
безусловные рефлексы, напротив, проявляются в процессе взаимодействия с другими
особями данного вида. Безусловные рефлексы саморазвития отражают
исследовательское поведение, рефлексы свободы, имитационный и игровой.
Нейрофизиолог Ю. М. Конорский
разделил безусловные рефлексы в соответствии с их биологической ролью',
сохранителъные, связанные с поступлением в организм и выведением из него
всего ненужного; восстановительные, направленные на сохранение вида
(копуляция, беременность, забота о потомстве), и защитные,
обеспечивающие удаление всего тела и его отдельных частей из сферы действия
опасного для организма раздражителя (рефлексы отдергивания и отступления) или
связанные с устранением вредящих агентов, которые попали на поверхность тела
или внутрь организма, уничтожением, а также нейтрализацией вредящих агентов
(наступательные рефлексы). Сохранительные рефлексы привлечения направлены
непосредственно на объект (пища, половой партнер), защитные направлены в
сторону, противоположную вредящему раздражителю. По порядку последовательности
фаз эта классификация дополняется указанием на подготовительные (мотивационные)
и исполнительные (консуматорные), связанные с завершающими действиями,
безусловные рефлексы. Так, исходя из этой классификации можно выделить подготовительные
пищевые безусловные рефлексы, лежащие в основе формирования состояний
голода и сытости. К ним относятся реакции, возникающие при изменении
химического состава крови, сдвигах в обмене веществ, усилении или ослаблении
интероцептивной сигнализации (преимущественно от рецепторов желудка, кишки и
печени).
Инициация и прекращение пищевого
возбуждения определяются нервными и гуморальными сигналами, воспринимаемыми
специализированными рецепторами гипоталамической области. В формирование
состояний голода и сытости вовлекаются также многие другие структуры головного
мозга. Пищевое побуждение зависит от внутренних раздражителей и от стимулов,
исходящих из внешней среды. На фоне доминирующей мотивации голода возникает
двигательное беспокойство и происходит активация некоторых сенсорных систем (в
частности, вкуса и обоняния). После поступления пищи в ротовую полость
подготовительные рефлексы тормозятся и начинают реализоваться исполни
тельные пищевые рефлексы: пережевывание пищи, слюноотделение, проглатывание
сформировавшегося пищевого комка, координированные сокращения пищевода и
желудка, выделение желудочного и панкреатического соков, изменение
метаболических реакций и т. д.
Столь же сложны подготовительные и
исполнительные безусловные рефлексы, связанные с половым или оборонительным
поведением. Следует вместе с тем иметь в виду, что в процессе онтогенеза
подготовительные и исполнительные безусловные рефлексы меняются под .влиянием
внешних и внутренних раздражителей, поэтому в координированной
приспособительной деятельности первостепенную роль начинают играть условные
рефлексы.
Как можно было видеть, рефлекторное
управление функциями организма осуществляется механизмами разной сложности. Это
позволило И. П. Павлову разделить безусловные рефлексы по анатомическому
принципу: простые
(спинномозговые), усложненные (продолговатый мозг), сложные
(средний мозг) и сложнейшие (ближайшие подкорковые образования и кора
больших полушарий). При этом И. П. Павлов указывал на системный характер
регуляции физиологических процессов, рассмотренный им на примере организации
«пищевого центра» — функциональной совокупности структур, расположенных на
разных уровнях головного мозга.
Положение о системности как
основном принципе работы мозга было сформулировано А. А. Ухтомским в его учении
о доминанте — функциональном объединении различных нервных центров на
основе повышенной возбудимости. Эти идеи были развиты П. К. Анохиным, согласно
представлениям которого функциональные системы динамически объединяют нервные
элементы различных уровней ЦНС, обеспечивая определенные приспособительные
эффекты.
Таким образом, существует
возможность классифицировать безусловнорефлекторную и условнорефлекторную
деятельность на основании анатомического и функционального подходов, между которыми
нет принципиальных противоречий. В последние десятилетия с помощью
стереотаксической техники удалось определить участие в специализированной
безусловнорефлекторной деятельности многих отделов мозга (гипоталамуса,
миндалевидного тела, гиппокампа, стриопаллидарной системы и др.). Полученные
данные расширили представления об организации различных форм поведения.
Развитие теории автоматического
регулирования привело к необходимости рассмотреть организацию врожденного и
приобретенного поведения в аспекте представлений об информационно—управляющей
деятельности мозга. Было выделено шесть уровней его организации:
элементарные, координационные, интегративные, сложнейшие безусловные рефлексы,
элементарные безусловные рефлексы и сложные формы высшей нервной (психической)
деятельности.
Элементарные безусловные рефлексы — простые ответные реакции местного
значения, реализуемые в соответствии с жестко детерминированной программой
своих сегментарных центров. Они осуществляются по одному главному каналу
(центростремительные, центральные и центробежные звенья). Роль обратных связей
(преимущественно отрицательных) в коррекции элементарных безусловных рефлексов
невелика. Примеры такого рефлекса — отдергивание обожженной руки от огня или
мигание при попадании в глаз соринки.
Координационные безусловные рефлексы также осуществляются на
сегментарном уровне, но в отличие от элементарных рефлексов включают ряд
циклов, хотя и стереотипных, но допускающих коррекцию на основе отрицательных и
положительных обратных связей. Пример простого координационного рефлекса —
антагонистический рефлекс, согласующий сокращения сгибательных и разгибательных
мышц.
Интегративные безусловные рефлексы — комплекс взаимно координированных
двигательных и вегетативных реакций, лежащих в основе биологически значимых
актов и способствующих поддержанию гомеостаза и коррекции элементарных
рефлексов. Реализация интегративных рефлексов определяется надсегментарными
механизмами (в основном нижними отделами ствола, структурами продолговатого,
среднего и промежуточного мозга, мозжечка). Если для осуществления элементарных
и координационных рефлексов имеют значение главным образом физические свойства
и локальное приложение раздражителя, то интегративные рефлексы обеспечивают
целостные ответы организма (простейшие поведенческие акты с их вегетативными
компонентами).
Механизмы нервной регуляции разных
уровней тесно переплетаются, поэтому их разделение носит условный характер.
Даже у спинального животного в осуществлении элементарного рефлекса участвует
несколько рефлекторных дуг. Еще И. М. Сеченов обнаружил, что у лягушки
неэффективность удаления лапкой повреждающего раздражителя приводит к
вовлечению в реакцию новых двигательных координации. Двигательный ответ
определяется исходным состоянием рефлекторного аппарата. У обезглавленной
лягушки раздражение кожи стопы вызывает ее сгибание, при согнутой — разгибание.
Нетрафаретность реализации врожденных рефлекторных программ, проявляемая даже
после удаления надсегментарных отделов центральной нервной системы, значительно
сильнее выражена при сохранении ее целостности.
Сложность организации врожденных
реакций
прослеживается на примере слюноотделительного безусловного рефлекса, который
принято было считать относительно простым. В действительности же он связан с
различными рецепторами (вкусовыми, тактильными, болевыми), волокнами нескольких
нервов (тройничного, лицевого, языкоглоточного, блуждающего), многими отделами
ЦНС (продолговатым мозгом, гипоталамусом, миндалевидным телом, корой больших
полушарий). Слюноотделение связано с пищевым поведением, сердечно—сосудистой,
дыхательной, эндокринной, терморегуляторной функциями.
Безусловнорефлекторная секреция
слюны зависит не только от вызывающего ее адекватного раздражителя, но и от
многих внешних и внутренних факторов. Повышение температуры окружающей среды
приводит у собак к выделению большого количества «терморегуляторной» слюны с
низким содержанием органических веществ. Количество слюны зависит от уровня
пищевого возбуждения, доступности и содержания в пище поваренной соли, гормонального
фона и многих других факторов.
Таким образом, реализация, казалось
бы, относительно простых врожденных реакций определяется системной интеграцией
сложных механизмов поддержания гомеостаза и взаимоотношений организма с внешней
средой. Такая интеграция чрезвычайно пластична, и, в соответствии с принципом
доминанты, одни и те же реакции могут входить в комплексы, связанные с
удовлетворением различных потребностей организма. Например, слюноотделительный
рефлекс может быть связан с терморегуляцией, пищевым или оборонительным
поведением.
В осуществлении интегративных
безусловных рефлексов, представляющих собой комплексы координированных движений
с их вегетативным обеспечением, ведущую роль играют надсегментарные механизмы.
Комплекс обратных связей осуществляет коррекцию элементарных, координационных и
интегративных реакций, объединяемых в единую систему. Она неотделима от
центральных механизмов инстинктивных реакций, связанных с подкорково—стволовыми
отделами головного мозга. Определенную роль в реализации инстинктивных реакций
играет также кора больших полушарий головного мозга.
Можно заметить, что предлагаемое
разными авторами разделение уровней безусловнорефлекторной деятельности
относительно. Схематичность любой ее классификации прослеживается на примере
одного из фундаментальных безусловных рефлексов — ориентировочного. Он
включает три группы явлений. Первая его форма, обозначенная И. П.
Павловым как рефлекс «что такое?», включает много элементарных и
координированных реакций — расширение зрачка, снижение порогов чувствительности
к ряду сенсорных раздражителей, сокращение и расслабление мышц глаза, уха,
поворот головы и туловища в сторону источника раздражения, принюхивание к нему,
изменение электрической активности головного мозга (угнетение, блокада α—ритма
и возникновение более частых колебаний), появление кожно—гальванической
реакции, углубление дыхания, расширение кровеносных сосудов головы и сужение
сосудов конечностей, начальное замедление и последующее учащение сердцебиений и
целый ряд других изменений в вегетативной сфере организма.
Вторая форма ориентировочного рефлекса связана
со специализированными поисковыми движениями и зависит от
мотивационно—потребностных характеристик, т. е. превалирующей доминанты, и от
внешних раздражителей.
Третья форма ориентировочного рефлекса
проявляется в виде исследовательской реакции, не обязательно связанной с
удовлетворением текущих потребностей организма, т. е. основанной на
«любопытстве».
В зарубежной литературе для описания
ориентировочного рефлекса используют психологические понятия — внимание,
установка в условиях ожидания раздражителя, реакция удивления, настораживания,
испуга, тревоги, бдительности. С точки зрения нейрофизиолога, ориентировочный
рефлекс — многокомпонентная неспецифическая реакция организма на
«новизну», направленная на повышение способности анализаторов дифференцировать
новое явление. Он характеризуется эффектом угасания и независимостью от
модальности и направления изменений стимула.
Ориентировочно—исследовательский
рефлекс является составной частью ориентировочно—исследовательского
поведения, которое, будучи врожденным, тем не менее практически неотделимо
от условнорефлекторной деятельности. Это касается и многих других форм
поведения. Поэтому одним из наиболее сложных вопросов физиологии поведения
является разделение врожденных и приобретенных реакций.
У взрослой особи врожденная
деятельность обычно не проявляется в чистом виде, она видоизменена
формирующимися в процессе онтогенеза условными рефлексами. Таким образом,
безусловные рефлексы модифицируются в соответствии с индивидуальным
приспособлением к особенностям существования. Даже на самых ранних этапах
постнатальной жизни, а для некоторых сторон жизнедеятельности еще в
пренатальном периоде, врожденные реакции «обрастают» условнорефлекторными
элементами. При этом генетически детерминированные положительные реакции могут
трансформироваться в отрицательные. Так, на самых ранних этапах жизни
предпочитаемый сладкий вкус может становиться отвергаемым, если он хотя бы
однократно сочетался с болезненным состоянием организма (дискомфортом).
Другая сложность в дифференцировании
врожденных и приобретенных реакций связана с совершенствованием
безусловнорефлекторной деятельности в процессе индивидуального развития.
Кроме того, при взаимодействии с условными рефлексами безусловные в процессе
постнатальной жизни «дозревают» (Л. А. Орбели).
Видоизменение врожденных форм
поведения в процессе индивидуального развития может зависеть не только от
научения, но и от многих косвенных влияний, в конечном счете
сказывающихся на безусловнорефлекторной деятельности. Она в ряде случаев
определяется температурой окружающей среды, при которой происходит развитие
организма, условиями питания, стрессорными воздействиями.
Поведение принято рассматривать как
врожденное, если в онтогенезе нельзя обнаружить влияний на него обучения или
других факторов. Эти влияния пытаются выявить при помощи экспериментов с
использованием определенных видов депривации (от англ. deprivation — лишение, утрата). (например, изоляции
от сверстников, выращивания в темноте и т. д.). Подобный метод не всегда
эффективен, поскольку депривация, во—первых, не может исключить всех средовых
воздействий, а во—вторых, вызывает ряд общих изменений в состоянии организма. В
частности, в зависимости от раздражителей, воздействующих на развивающийся
организм (обогащенная и обедненная среда), регулируются синтез ДНК в нейронах,
нейромедиаторный баланс и многие другие составляющие, от которых зависит
реализация поведенческих актов.
Ответы организма не являются
следствием прямолинейных процессов развития, ведущих от гена прямо к поведению
взрослого животного и лишь в некоторых случаях изменяемых внешними
воздействиями. В действительности существует сложное переплетение причинных
связей, когда каждая часть организма может взаимодействовать с другими его
частями и внешней средой.
Диапазон изменчивости сложнейших
безусловных рефлексов в зависимости от условий существования в раннем возрасте
неодинаков для разных видов деятельности. Некоторые врожденные комплексы
движений чрезвычайно устойчивы и не могут быть изменены воздействиями среды,
другие отличаются большей пластичностью. Описаны фиксированные
последовательности движений, не зависящие от научения. Они хорошо
прослеживаются у насекомых и птиц. Так, роящиеся осы одного вида строят гнезда
при помощи стереотипных движений, шаблонны движения домашних петухов при
ухаживании за курами.
Фиксированные комплексы движений
характерны и для высокоразвитых животных, в том числе и для человека.
Характерны сканирующие движения головой младенцев, облегчающие поиск соска.
Стереотипно проявляются другие комплексы движений, связанные с сосанием. Эти
рефлексы созревают еще в пренатальном периоде развития, что установлено в
наблюдениях над недоношенными детьми. Не зависят от научения хватательный
рефлекс, мимика ребенка и многие другие проявления врожденной деятельности.
Наблюдения над представителями многих видов животных показывают, что адекватный
выбор пищи может осуществляться без помощи родителей, т. е. не всегда требует
предварительного научения. Отрицательная реакция на высоту проявляется у
обезьянок, выросших в зоопарке.
Вместе с тем многие сложнейшие
безусловные рефлексы видоизменяются в процессе развития или же требуют для
своего проявления периода научения. У птенцов формирование пения определяется
не только врожденными характеристиками, но и условиями вскармливания птицами
своего или другого вида. Изоляция от сверстников крысят или щенков приводит к
необратимым изменениям последующего «социального» общения. Изоляция обезьянок
резко нарушает их последующее половое и материнское поведение.
Трудности, возникающие при
разделении детерминированных генетически и выработанных в процессе жизни
поведенческих актов, усугубляются и тем обстоятельством, что некоторые
врожденные формы поведения проявляются на относительно поздних этапах развития,
когда животное обладает некоторым опытом и уже сформированы условнорефлекторные
стереотипы.
Так происходит, в частности, с половым
поведением, готовность к проявлению которого возникает в определенном
возрасте на фоне гормональных перестроек. Однако эффективность спаривания у
многих видов определяется также индивидуальным опытом, приобретаемым до
достижения половой зрелости в результате общения со сверстниками. Например, у
взрослых самцов цихловых рыб, выращенных в изоляции, поведение ухаживания
адресуется не только самкам, но и самцам. Близкие изменения прослежены у птиц,
грызунов, обезьян. Общение с сородичами влияет на половое поведение разными
путями, изменяя готовность к спариванию, реактивность к соответствующим
раздражителям, точность движений и различные реакции, прямо или косвенно
связанные с размножением. При этом следует иметь в виду, что специфическое (в
данном примере половое) поведение может видоизменяться у взрослых особей на
основе неспецифического по отношению к нему поведения, проявляющегося на более
ранних стадиях онтогенеза.
Половое созревание и связанные с ним
гормональные перестройки изменяют многие стороны взаимодействия с внешней
средой, непосредственно не относящиеся к репродуктивным процессам. У взрослых
особей половые различия проявляются в двигательной активности, поведении в
открытом поле, внутривидовой агрессии, в восприятии вкусовых, обонятельных и
болевых раздражителей, выборе и потреблении пищи, в научении пассивному и
активному избеганию повреждающего воздействия, научении в лабиринте. По мере
полового созревания изменяется характер реагирования на многие биологически
значимые раздражители, что, в свою очередь, отражается на реализации ранее
выработанных условных рефлексов. Эта закономерность хорошо прослежена на
примере условнорефлекторных вкусовых аверсий — отрицательного отношения
к врожденно индифферентным или предпочитаемым вкусовым стимулам после сочетания
их с болезненным состоянием. Так, аверсия к сладкому вкусу, однократно
сочетавшемуся с отравлением, одинаково выражена у неполовозрелых крысят обоего
пола. По мере полового созревания у самок увеличивается связанная с повышением
уровня эстрогенов и прогестинов мотивация к потреблению веществ сладкого вкуса
и, соответственно, снижается выработанная к ним аверсия. У самцов же она
сохраняется значительно дольше, что поддерживается продукцией тестостерона.
После кастрации вкусовые аверсий у самцов угашаются так же быстро, как и у
самок.
Не следует, однако, связывать
половой диморфизм (морфофизиологические различия между особями мужского и
женского пола) в различных формах поведения, в том Числе и с вкусовыми
аверсиями, исключительно с состоянием эндокринной системы. Он определяется
совокупностью факторов, связанных с особенностями формирования центральной
нервной системы в пре— и постнатальном онтогенезе, гормональным статусом
организма и возможностью реализации тех или других форм активности в
соответствии с внешними условиями. Имеется, например, корреляция между
пролонгированием сохранения условнорефлекторных вкусовых аверсий и
пассивно—оборонительным поведением. Очевидно, более высокий уровень последнего
у самцов по сравнению с самками обеспечивает надежность защиты «гнезда».
Анализ проявлений полового диморфизма
целесообразно проводить с позиций представления о стратегиях и тактиках
поведения на разных этапах индивидуального развития (В. Г. Кассиль).
Преимущественный выбор активных и пассивных форм реагирования на экстремальные
раздражители определяется отношением к ситуации как избегаемой или
неизбегаемой, нередко не совпадающим с ее реальной оценкой, что обусловлено
предшествующим опытом. Вместе с тем исходная направленность реагирования на
внешние события обусловлена врожденными характеристиками и в значительной мере
зависит от особенностей организации мозга и гуморально—гормонального статуса
организма. Так, анализ диморфизма в поведении самцов и самок показывает, что у
последних преобладают активные формы реагирования на экстремальные
раздражители.
Для проявления различных врожденных
форм поведения и вырабатываемой на их основе условнорефлекторной деятельности
чрезвычайно важны процессы созревания центральной нервной системы и
сопутствующие изменения в балансе биологически активных веществ во внутренней
среде. С этими процессами тесно связаны особенности взаимодействия безусловно—
и условнорефлекторной деятельности на разных этапах онтогенеза. Существенными
для раннего взаимодействия со средой являются реакции, проявляющиеся в первые
часы или дни жизни у птиц и млекопитающих при однократном сочетании раздражения
различных органов чувств с врожденными элементами поведения — следованием за
удаляющимся предметом и с другими целенаправленными двигательными актами.
Эта форма обучения, названная запечатлением
(импринтингом), формируется на протяжении чувствительного периода,
длящегося от 6—8 ч до 4—5 дней. Близки к запечатлению натуральные условные
рефлексы, которые также формируются очень быстро на определенной стадии
онтогенетического развития и чрезвычайно медленно угасают. Можно выделить форму
научения, имеющую черты импринтинга и натурального условного рефлекса. Так, у
щенков на протяжении первых трех дней жизни вырабатывается пищедобывательный
условный рефлекс на естественные или экологически неадекватные запаховые
раздражители при однократном их сочетании с кормлением. С 4—го по 10—й день
жизни способность к выработке этого рефлекса исчезает и вновь появляется на 11
~12—й день, причем начиная с этого периода научение требует уже многократного
сочетания условного и безусловного стимулов.
Сложные формы поведения проявляются сразу же после
рождения, что позволяет отнести их к врожденным реакциям. Процесс их дозревания
не может быть прослежен «в чистом виде», так как они модифицируются благодаря
внешним воздействиям. Существование феноменов запечатления и натуральных
условных рефлексов затрудняет дифференцирование врожденных реакций,
приобретенных в постнатальном онтогенезе поведенческих актов.
Есть основания предполагать, что
реализация некоторых врожденных реакций связана с раздражителями, которым
подвержен организм в пренатальной жизни. Так, у щенков предпочтение запаха
матери формируется в конце пренатального периода.
Некоторые врожденные реакции
проявляются не сразу после рождения, а на одном из последующих этапов развития.
Если в это время животное не сталкивается со специфическим раздражителем, в
дальнейшем способность реагировать на него без специального научения не
проявляется. При этом возможны ошибки в отнесении некоторых реакций к
врожденным или выработанным. Например, долгое время считали, что собаки,
воспитывавшиеся с периода перехода к дефинитивному питанию на хлебно—молочной
диете, не отвечают врожденной положительной реакцией на запах мяса. Первые
опыты на этих животных были проведены лишь в 7—месячном возрасте. Оказалось,
однако, что на 16—21—й день жизни щенка такая способность у него проявляется.
Если же адекватный раздражитель отсутствует, она постепенно затормаживается и
отсутствует у щенков старшего возраста, впервые сталкивающихся с запахом мяса.
Проявление некоторых сложных форм
поведения, хотя и определяется генетической программой, в некоторых пределах
может модулироваться внешними факторами. Так, понижение температуры внешней
среды значительно снижает уровень игровой деятельности детенышей некоторых
млекопитающих, хотя вызывает ее специфический раздражитель — контакт со
сверстниками.
Можно привести много примеров,
подтверждающих роль факторов среды в модификации врожденных форм поведения.
Однако было бы ошибкой противопоставлять значение генетических и средовых
факторов в развитии поведения. Все формы взаимодействия организма со
средой, в том числе и поведенческие, обусловлены генетической программой и в
той или иной степени подвержены внешним влияниям. Генетическая программа
определяет и диапазон этих влияний, т. е. так называемую норму реакции.
Для одних признаков она строго фиксирована, что хорошо иллюстрирует отсутствие
пластичности в осуществлении некоторых функций у насекомых (полет, выход из
личинки или кокона, половое поведение).
Существуют строго
запрограммированные инстинктивные действия. Например, самка паука при
строительстве кокона производит комплексы стереотипных движений, даже если
паутинная нить не вырабатывается. Затем в несуществующее отверстие она
откладывает яйца, которые падают на землю, и продолжает деятельность,
имитирующую строительство кокона, которого на самом деле нет. В этом случае
норма реакции крайне узка и инстинктивные действия не зависят от сигналов об их
эффективности. Для ряда других признаков она значительно шире, причем
адаптивная вариабельность инстинктивных действий обнаружена и у насекомых, что
проявляется, в частности, при восстановлении разрушенных жилищ в условиях,
отличных от естественных.
Генетическая обусловленность
поведения проявляется при постепенном формировании некоторых поведенческих
актов в процессе раннего онтогенеза. Подробно исследовано соотношение
врожденных и приобретенных компонентов в реакции нападения на жертву у котят.
Вначале проявляются только инстинктивные Двигательные стереотипы, постепенно в
процессе тренировки, происходящей в условиях контакта с матерью и сверстниками,
они уточняются и обогащаются движениями, сформировавшимися в процессе научения.
Исходное использование врожденных
комплексов движений, связанных с пищевой активностью, описано у щенков первых
дней жизни при выработке внутримозговой самостимуляции «зон награды»
(положительной эмоциогенной системы). Постепенно репертуар движений обогащается
менее шаблонными, выработанными комплексами, причем они соседствуют с
врожденными стереотипами двигательной активности. Очевидно, инстинктивные
поведенческие акты, на основе которых строится новая система целенаправленной
активности, не обязательно устраняются при ее формировании.
Сложным является вопрос о
непременной рефлекторной основе каждого поведенческого акта.
Представление о ее обязательности
привело И. П. Павлова к отождествлению понятий сложные безусловные реакции
и инстинкты. Во многих случаях удалось обнаружить внешние и внутренние
стимулы, являющиеся толчком к развертыванию цепи шаблонных реакций, однако не
всегда можно их идентифицировать, что позволяет предполагать, что ряд форм
инстинктивной деятельности проявляется самопроизвольно. Эндогенные процессы
в ЦНС обусловливают выполнение ряда инстинктивных актов без видимых колебаний в
состоянии внешней и внутренней среды. Большую роль при этом играют циркадные
(околосуточные) и другие ритмы, которые не определяются физиологическим
состоянием организма и различным стимулами, хотя и могут сдвигаться под их
влиянием.
Описаны
автономные колебательные процессы в различных структурах мозга, которые
определяют периодические изменения поведения животных, изолированных от
сородичей и лишенных зрения и слуха. Многие генетически закодированные реакции
определяются изменениями во внутренней среде организма. Так, у глухих от
рождения сиамских кошек—мутантов возбуждение, связанное с циклом половой
активности, проявляется в поведенческих актах (лордоз — прогибание
позвоночника, обращенное выпуклостью вентрально, и др.) ив специфических
звуковых сигналах. Определенные сигналы издаются этими животными в состоянии
голода и при оборонительном поведении.
Некоторые нормы реакций при
отсутствии обратной связи подавляются. Так у глухих и слепых людей отсутствуют
некоторые выразительные движения (в том числе и звуковые), связанные
соответственно со слуховым или зрительным восприятием. Рожденные слепыми с
годами улыбаются меньше, чем зрячие или ослепшие в более позднем возрасте.
Однако ряд выразительных движений проявляется независимо от полноценности сенсорных
систем. Анализ зафиксированных на кинопленку выразительных движений детей,
родившихся слепыми и глухими, показал, что моторика смеха у них точно такая же,
как и у здоровых.
Комплексы инстинктивных движений
обычно тесно связаны с сигналами из внешней и внутренней сред организма, хотя
они и могут определяться автономными процессами в ЦНС. Однако далеко не всегда
удается их выявить.
Отрицание рефлекторной природы
инстинктивных актов
привело некоторых исследователей к определению их как врожденных, внутренне
организованных и проявляющихся спонтанно. Полагают, что инстинкт связан с
накоплением «специфической энергии действия», которую освобождает разрешающая
ситуация. При этом инстинктивные действия, отражающие внутренние потребности,
включают поисковую (подготовительную) и завершающую фазы.
Примером может служить активность
хищника при выслеживании и поедании жертвы. На первом этапе имеется
ненаправленный поиск, далее в соответствии с раздражителями, исходящими от жертвы,
поиск становится направленным, после чего следует ряд поведенческих актов
(подкрадывание или преследование, прыжок, умерщвление жертвы, расчленение ее на
куски). Второй этап (поедание жертвы) является завершающим (консуматорным) и
протекает более стереотипно, чем первый.
3.13.2. Достижения этологов в исследовании врожденных форм
поведения
Несмотря на терминологические
различия в обозначении некоторых поведенческих реакций, этологи описывают их с
позиций, весьма близких позициям И. М. Сеченова, И. П. Павлова и других
отечественных физиологов.
В чем же заключаются основные
положения современной этологии? Согласно концепции К. Лоренца,
инстинктивное поведение состоит из комплексов фиксированных паттернов —
одинаковых у всех особей данного вида стереотипных по последовательности и
форме исполнения двигательных актов. Под действием внутренних и внешних
раздражителей (гормонального фона, температуры внешней среды и др.) в нервных
центрах накапливается «специфическая энергия действия». Она вызывает поисковое
поведение, направленное на удовлетворение определенного побуждения (голода,
жажды, половой потребности и т. п.).
Фаза поиска включает не только врожденные, но и
приобретенные элементы тогда как следующая за ней завершающая фаза, которую
можно считать инстинктом в чистом виде, строго видоспецифична и
модифицируется в процессе научения. Можно привести множество примеров
шаблонности завершающей фазы поведенческого акта, на основании которых были
расширены возможности систематики. При использовании в качестве таксономических
признаков комплексов фиксированных действий завершающей фазы поведенческого
акта удалось дифференцировать виды, не различающиеся морфологически.
Так, 16 видов уток были разделены на
основании комплексов фиксированных действий при церемонии ухаживания.
Существенные коррективы были внесены в классификацию семейства кошачьих, у
которых формирование охотничьих навыков и типов общения были сопоставлены с
анатомическими и экологическими особенностями.
Согласно концепции этологов, центральная
нервная система блокирует комплексы фиксированных действий. Блок снимается
ключевыми, или знаковыми, раздражителями (стимул—объекты, рилизеры),
высвобождающими «врожденный разрешающий механизм». Избирательный ответ на эти
раздражители связан с нейросенсорными системами, запускающими данную реакцию и
определяющими избирательную чувствительность к специфическим комбинациям
ключевых стимулов. Отношение к ним меняется в зависимости от внутреннего
состояния. Чайки, например, могут относиться к яйцам как к пище или объекту
высиживания. После освобождения из куколки самец бабочки—сатира сильнее
реагирует на желтый и синий цвета — ключевые стимулы, облегчающие поиск цветка.
В период же ухаживания ключевым стимулом становится коричневый цвет.
Ключевые стимулы исследуют с помощью
макетов, в которых последовательно исключают детали естественного
носителя ключевых раздражителей, а также изменяют его размеры. Можно изготовить
«сверхнормальную» модель, в которой выделен ключевой раздражитель. Животные
предпочитают такую модель естественному комплексу раздражителей.
В опытах, проведенных на серебристой
чайке, сером гусе и кулике—сороке, обнаружено, что для насиживания птицы
выбирают огромные макеты яиц, предпочитая их собственным. Птенцы серебристой
чайки клюют макет клюва взрослой чайки (врожденная реакция, вызывающая
отрыгивание родителями рыбы), который привлекает их больше, чем естественный,
хотя значительно от него отличается. Наибольшую реакцию вызывает тонкая белая
палочка с поперечными темно—красными полосками. У самца колюшки во время
брачного сезона брюшко окрашивается в ярко—красный цвет, что провоцирует у
другого самца агрессивную реакцию. Она может быть вызвана любым продолговатым
предметом, имеющим снизу красное пятно. Все эти стимулы являются врожденными и
не корригируются индивидуальным опытом.
Видоспецифические зрительные
раздражители охватывают обширную категорию сигналов, определяющих различные
формы инстинктивного поведения. Однако они составляют лишь часть ключевых
стимулов, которые могут быть адресованы различным сенсорным системам. Хорошо
исследованы звуковые сигналы, связанные с пищевым, оборонительным, половым
поведением, взаимоотношениями матери и детеныша, членов сообщества
(зоосоциальное поведение), Пусковую роль для различных форм врожденной
активности играют феромоны — вещества, вырабатываемые специальными
железами или клетками животных, выделяемые в окружающую среду и имеющие
сигнальное значение.
Ключевые
стимулы, как правило, относительно просты. Между тем общение животных включает
комплексы сложных фиксированных движений, которые могут быть ключевыми
стимулами для разных форм поведения. Хорошо изучена передача информации,
касающейся источников пищи у пчел. У представителей различных классов животных
описаны позы угрозы и подчинения, ритуалы приветствий, брачные церемонии.
Демонстративные движения часто отражают конфликтные тенденции к двум
несовместимым типам поведения, например, оборонительному и половому.. В
зависимости от уровня мотивации и возможности ее реализации побеждает одна из
таких тенденций. Если конфликт не может разрешиться, между конкурентными
мотивациями на какое—то время устанавливается равновесие. В этом случае может
проявиться смещенная активность — деятельность, не соответствующая
исходной ситуации. Так, вместо проявления агрессивного или полового поведения
птицы начинают чистить клюв
или оперение, клевать землю или зерна (часто не глотая их). Смещенная
активность изучена у многих представителей птиц, рыб, членистоногих,
млекопитающих. В состоянии напряжения люди также бессознательно совершают
ненужные в данной ситуации действия, например поправляют галстук,
причесываются. Смещенная активность может зависеть от внешних стимулов. У
индюков в конфликтной ситуации «приближения—избегания» она проявляется в
потреблении пищи или воды в зависимости от их доступности. Внутреннее состояние
также влияет на характер смещенной активности. Одно из частых ее проявлений у
птиц — чистка перьев — может инициироваться взъерошиванием, вызванным
конфликтной ситуацией.
Конфликт может быть разрешен с
помощью переадресованной активности — нападением на более слабую особь,
когда агрессия тормозится страхом.
Н. Тинберген выдвинул предположение
об иерархической организации инстинктивного поведения, проявляющейся в
последовательном включении отдельных комплексов движений в результате
упорядоченного возбуждения соответствующих «центров».
Сопоставление подходов к изучению
инстинктивной деятельности, характерной для этологов и зоопсихологов,
показывает, что они обращают недостаточно внимания на центральную организацию
поведения. Практически многие положения этологов, например представление о
специфической энергии действия иерархии центров, врожденном реализующем
механизме, реафферентации, далеки от физиологического подхода к изучению
врожденного и приобретенного поведения. Между тем учение И. П. Павлова о высшей
нервной деятельности, А. А. Ухтомского о доминанте, многие положения теории
функциональных систем П. К. Анохина, а также результаты исследований ряда
физиологов вскрывают конкретные механизмы врожденных форм поведения, описанных
этологами. Однако следует отдать должное огромной работе, проделанной
последними по наблюдению за различными сторонами поведения животных.
3.14. ПРИОБРЕТЕННЫЕ ФОРМЫ ПОВЕДЕНИЯ
Жизнедеятельность организма в
постоянно меняющейся внешней среде включает в качестве важнейшего фактора
широкий спектр индивидуальных поведенческих адаптации. Генетически
детерминированные формы поведения, отражающие накопленный в генофонде видовый
опыт предшествующих поколений, оказываются недостаточными, чтобы обеспечить
активное существование особи в вероятностно изменчивой среде. Чем меньше
выражен диапазон колебаний факторов среды, тем в большей степени поведение
животного может опираться на опыт предшествующих поколений, «записанный» в его
генотипе. И наоборот, чем больше изменчивость окружающей действительности, тем
меньше прагматическая ценность видового опыта, тем в большей мере возрастает
необходимость в приобретении собственного, индивидуального опыта.
Индивидуальный опыт приобретается
различными путями, в основе которых лежит общая способность живых организмов к
научению, что возможно благодаря важнейшему свойству организма сохранять на
какой—то срок элементы научения, т. е. свойств памяти.
Нейробиология научения и памяти —
это единая научная проблема. Под научением понимают процесс, состоящий в
появлении адаптивных изменений индивидуального поведения в результате
приобретения опыта. Этот процесс может быть оценен по результатам поведения,
иными словами, по тем поведенческим актам, которые животные запомнили в
результате научения.
3.14.1. Классификация форм научения
В основу классификации, которая
опирается на ранее разработанные схемы, положены филогенетическая и
онтогенетическая динамика развития живого организма.
Молодые организмы с первых моментов
самостоятельной жизни обеспечены достаточно стабильным набором внешних
стимулов, среди которых они находятся. На этом этапе научение носит неассоциативный
облигатный (истинный) характер, обусловленный набором средовых факторов и не
требующий их непременной ассоциации с целостной деятельностью организма.
Научение имеет стимулзависимый характер.
К этой форме научения могут
относиться суммационный рефлекс, привыкание, запечатление (импринтинг),
подражание (имитация). В основе феномена суммации лежат явления сенсибилизации
(повышение чувствительности организма к данным раздражающим агентам) и фасилитации
(облегчение запуска именно данной ответной реакции). Такие поведенческие навыки
описаны даже у одноклеточных простейших и беспозвоночных. Это — освоение
определенного маршрута передвижения, скопление в определенных местах
пространства, различение съедобных продуктов и несъедобных и пр.
Эти и подобные им навыки обладают
следующими свойствами: 1) они не могут сохраняться длительное время и обречены
на исчезновение; 2) вызывающие их раздражители не имеют специализированного
сигнального значения;
3) после
исчезновения эти навыки самостоятельно не восстанавливаются.
Последнее свидетельствует о том, что
исчезновение этих навыков есть процесс разрушения, а не временного торможения.
Возникнув на ранних этапах эволюции живых существ, суммационная реакция у
высших животных в преобразованном, а иногда и замаскированном виде проявляется
как важнейший элемент более сложных форм индивидуального научения.
В противоположность суммационной
реакции привыкание представляет собой такую форму научения, которая состоит в
относительно устойчивом ослаблении реакции вследствие многократного
предъявления раздражителя, не сопровождающегося каким—либо биологически
значимым агентом (пищевым, оборонительным и пр.).
По—видимому, для низкоорганизованных
животных суммационная реакция и кратковременное привыкание являются единственными
механизмами приобретение индивидуального опыта. Привыкание совершенствуется
в ходе эволюции и сохраняет за собой важную роль в репертуаре поведенческих
адаптации. В определенном смысле привыкание — это подавление реакций,
значимость которых невелика для поддержания жизнедеятельности. Привыкание
возникает не в результате утомления или сенсорной адаптации, оно может быть
кратко— и долговременным, что позволило использовать привыкание как модель для
изучения разных видов памяти.
Наиболее распространенной формой
привыкания в животном мире является ориентировочный рефлекс, который при
повторении вызвавшего его раздражителя угашается в соответствии со всеми
признаками привыкания. Основными факторами в развитии ориентировочного рефлекса
являются новизна, неожиданность и значимость раздражителя для организма. В составе
ориентировочной реакции выделяют два процесса: 1) начальную реакцию тревоги,
удивления, сопровождающуюся повышением тонуса мышц и фиксированием позы
(затаивание), генерализованным изменением электрической активности мозга
(неспецифическая настройка), и 2) исследовательскую реакцию внимания —поворот
головы, глаз, ориентация рецепторов по направлению к источнику раздражения. Она
сопровождается локальными изменениями электрической активности мозга.
Ориентировочная реакция возникает не
на сам стимул как таковой, а в результате сличения стимула со следовым
процессом, оставленным в нервной системе предшествующими раздражителями. Если
стимул и след совпадают, то ориентировочная реакция не возникает. Если же
информация о стимуле и хранящиеся в памяти следы не совпадают, ориентировочная
реакция возникает и оказывается тем интенсивнее, чем больше выражено такое
рассогласование. Вероятно, конфигурация следов, оставленных в нервной системе в
результате повторения каких—то раздражителей, фиксирует все их параметры.
Иными словами, нервная система путем
комбинации своих элементов строит «нервную модель» стимула, динамика
формирования которой и отражается в процессах угашения ориентировочного
рефлекса.
Первые шаги в самостоятельной жизни
молодого организма в решающей мере зависят от его контактов с матерью, которые
осуществляют его выживание. Комплекс поведенческих адаптации новорожденного,
которые образуют первичную связь между ним и родителями, как бы замыкает цепь
преобразований эмбрионального периода, позволяя реализовать новорожденному уже
сформированные механизмы восприятия и реагирования, именуют запечатлением
(импринтингом). Детальная биологическая характеристика этого явления
проведена К. Лоренцем, который сформулировал четыре основных особенности
импринтинга: 1) импринтинг приурочен к ограниченному отрезку жизни, именуемому критическим,
или чувствительным, периодом; 2) импринтинг необратим, т. е., возникнув
в критический период, он не уничтожается последующим жизненным опытом; 3)
импринтинг может происходить в тот период, когда соответствующее поведение еще
не развито (например, половое); т. е. научение путем импринтирования не требует
подкрепления; 4) при импринтинге запечатлеваются не индивидуальные, а
видоспецифические характеристики жизненно важного объекта.
Типичный пример импринтинга: если
утенок вылупляется в присутствии большого зеленого ящика, он будет следовать,
за ним, двигающимся по проволоке, причем даже более охотно, чем за собственной
матерью или другими птенцами. По—видимому, явления импринтинга служат примером
взаимодействия индивидуального опыта и врожденных свойств молодого организма,
что ведет к быстрой фиксации их в механизмах памяти.
В результате подражания
(имитации) животное выполняет видотипичные действия путем непосредственного
наблюдения за действием других животных своего вида. Особенно это характерно
для молодых животных, которые путем имитации родительского поведения научаются
различным проявлениям поведенческого репертуара своего вида.
Л. А. Орбели считал имитационное
поведение главным охранителем вида, ибо громадное преимущество
заключается в том, что «зрители», присутствующие при акте повреждения члена их
же стада или их сообщества, вырабатывают рефлекторные защитные акты и таким
образом могут в будущем избежать опасности.
Таким образом, облигатное,
неассоциативное научение обеспечивает жизнедеятельность особи на первых этапах
ее самостоятельного существования, а также закладывает основы
видоспецифического характера поведения.
На более поздних этапах онтогенеза
поведение все в большей мере приобретает активный характер. Расширяется спектр
внешних факторов, способных приобретать сигнальное значение в зависимости от
сочетания их с целостной биологической реакцией организма (ассоциативное,
факультативное научение). Научение в этот период носит эффектзависимый
характер, т. е. определяется результативностью контакта организма со
средой.
Ассоциативное научение
характеризуется совпадением во времени какого—либо индифферентного раздражителя
с деятельностью организма. Биологический смысл такой ассоциации — условного
рефлекса — в его сигнальности, т. е. в приобретении этим раздражителем
роли предупреждающего фактора, сигнализирующего наступление последующих событий
и подготавливающего организм к взаимодействию с ними. Ситуация выработки
условных рефлексов, как бы имитирующих безусловнорефлекторную реакцию,
наблюдалась в случае слюноотделительных или электрооборонительных рефлексов.
В последнем случае, например,
посторонний раздражитель сочетали со слабым электроболевым стимулом, приводящим
к отдергиванию конечности' у собаки. Спустя несколько сочетаний эта же реакция
возникала и на условный сигнал. Такие условные рефлексы, благодаря которым
достигается пассивная адаптация за счет существенного расширения сигналов к
безусловнорефлекторной деятельности, называют условными рефлексами I типа,
или классическими условными рефлексами. Животное при этом выступает в
качестве пассивного участника событий, не имеющего возможности кардинально
изменить их последовательность.
В самостоятельную форму объединяют
условные рефлексы, которые строятся на основе активной целенаправленной
деятельности животного. Теперь уже последовательность событий зависит не только
от внешней сигнализации, но и от поведения животного. Например, крыса в
экспериментальной камере случайно нажимает лапой на педаль, за что сразу
вознаграждается пищей. А если еще перед нажатием на педаль включать посторонний
раздражитель, устанавливается следующая цепная связь: сигнал →
нажатие на педаль → пища.
Активный характер такого научения
базируется на среднем звене — нажатии на педаль, от своевременности выполнения
которого зависит успешность выполнения пищедобывательного поведенческого акта.
Причем само нажатие на педаль не имеет никакой генетической связи с получением
пищи. Такие условные рефлексы именуют инструментальными, или условными
рефлексами II типа. Сюда же относят различные формы так называемого оперантного
обучения, или дрессировки, обучения с помощью «проб и ошибок».
Высшие формы научения, свойственные
в большей степени взрослым животным с развитой нервной системой, опираются на
свойство формировать функциональную структуру окружающей среды в концептуальном
плане, т. е. формировать целостный образ окружающей среды. Такие формы когнитивного
научения основаны на извлечении законов связей между отдельными
компонентами среды и базируются на двух предыдущих формах научения.
У высших
позвоночных животных при первом же восприятии местоположения пищи создается
образ или конкретное представление пищи и ее местоположения в данном
пространстве. Этот образ сохраняется длительное время без повторного
воспроизведения, и каждый раз, когда он репродуцируется при восприятии данной
среды или какого—либо ее компонента, животное ведет себя точно так же, как и
при непосредственном восприятии. Такое психонервное поведение, или поведение,
направляемое образами, стали называть произвольным в отличие от
условнорефлекторного, автоматизированного. При такой форме научения
устанавливаются временные нервные связи между психонервным образом и
двигательными центрами мозга. Образ, который лежит в основе субъективного
отражения внешнего мира, — это вектор предстоящего рефлекторного поведения.
Непосредственно активность образа реализуется через ориентировочную реакцию,
которая регулирует направление поведения животного. Индивидуальное поведение,
первично направляемое психонервным образом, при повторной тренировке
автоматизируется и осуществляется по всем закономерностям условнорефлекторного
научения.
В качестве самостоятельной формы
поведения выделяют элементарную рассудочную деятельность животных,
которая заключается в их способности улавливать простейшие эмпирические законы,
связывающие предметы и явления окружающей среды, и в возможности оперировать
этими законами при построении программ поведения в новых ситуациях.
Элементарным проявлением рассудочной деятельности (разума) животных является
способность к экстраполяции направления движения раздражителя. Животное,
используя свою «систему отсчета», экстраполирует направление смещения кормушки
с пищей, которая большую часть пути остается невидимой для животного. Таким
путем животное использует предварительно воспринятую тактику поведения в среде
для построения логики своего будущего поведения.
Предполагают,
что в процессе отбора закрепляются те нейронные функциональные констелляции,
которые обеспечивали наиболее быстрое решение биологически значимых задач.
Происходит сличение уловленных в данный момент законов с теми, которые были
познаны в предшествующей жизни. В результате осуществляется выбор наиболее
адекватного пути решения задачи. К проявлениям рассудочной деятельности следует
относить и так называемое инсайт—обучение (от англ. insight — проницательность, понимание,
интуиция.)
Важнейшим результатом биологической
эволюции в вероятностно организованной среде является способность к вероятностному
прогнозированию. Под этим понимают предвосхищение будущего, основанное на
вероятностной структуре прошлого опыта и информации о наличной ситуации. В
соответствии с вероятностными гипотезами осуществляется подготовка к действиям
в предстоящей ситуации, приводящим к наибольшей вероятности достижения цели.
Теория вероятностного обучения
исходила из представлений о предсказании статистических закономерностей и
выбора оптимальных стратегий поведения при обучении субъекта независимым
вероятностям стохастично подаваемых раздражении. Степень информированности
субъекта зависит от текущей субъективной вероятности достижения цели с учетом
наличных средств и оценки проблемной ситуации. Высшие позвоночные и человек
непрерывно опираются на вероятностное прогнозирование. Животные прогнозируют
вероятность поиска пищевого объекта, чтобы не стать жертвой собственной
«беспечности».
В целом вероятностное
прогнозирование может иметь разные формы: 1) прогнозирование разных форм не
зависимых от субъекта событий; 2) прогнозирование своих ответных активных
действий; 3) прогнозирование целенаправленных действий не только в соответствии
с их частотой в прошлом опыте, но и с их актуальной значимостью и
предполагаемым результатом; 4) использование гипотез о наиболее вероятных
действиях своих активных партнеров; 5) прогнозирование действий и целей с
учетом собственных энергетических затрат.
Обучение вероятностному
прогнозированию деятельности при широком репертуаре простых форм обучения
представляет собой результат наиболее сложных мозговых процессов, связанных с
работой высших интегративных систем мозга.
3.14.2. Сон как форма приобретенного поведения
(В изложении раздела использованы
материалы Ю. Ф. Пастухова).
Цикличность всех природных процессов
связана с движением Земли вокруг Солнца, с вращением Земли вокруг собственной
оси. Первое определяет смену сезонов года и соответствующие сезонные ритмы
поведения, последнее — смену дня и ночи, т. е. суточные, или циркадные,
ритмы поведения. Большинство млекопитающих не рождаются с готовым суточным
ритмом, а постепенно приспосабливаются к нему, т. е. вырабатывают собственный
внутренний ритм. Чем быстрее и полнее животное его сформирует, тем оно имеет
больше шансов в борьбе за существование. Периодическая смена освещенности и
сопутствующих внешних раздражении (прежде всего температурных) играет особо
важную роль в период становления внутреннего ритма. Эти ритмы могут быть
устойчивыми и поддерживаться без непосредственного сопровождения факторами
внешней среды. Смена сна и бодрствования — это одно из поведенческих проявлений
внутреннего суточного ритма организма.
Сон — это адаптация, подавляющая
активность в период наименьшей доступности пищи, угрозы резких колебаний
внешних условий и максимальной опасности со стороны хищников. С этими же
факторами связана общая продолжительность и структура сна. Сон снижает
ежедневные метаболические затраты, навязывая организму смену деятельности, и
является выражением внутренней инстинктивной потребности.
Сон бывает двух типов: монофазный
— с однократным чередованием дневного или ночного сна и периода бодрствования, полифазный
— с частыми сменами периодов сна и бодрствования на протяжении суток. По
современным представлениям, весь период одного цикла сна разделен на два сменяющие
друг друга и резко различающиеся состояния, которые именуются медленноволновым,
или медленным (ортодоксальным), сном (МВС) и быстрым, или парадоксальным,
сном (ПС). Эти названия связаны с тем, что стадия медленного сна
сопровождается высокоамплитудными медленными δ—волнами ЭЭГ, а стадия
быстрого сна — высокочастотной низкоамплитудной активностью (десинхронизация),
столь характерной для ЭЭГ мозга бодрствующего организма. Таким образом, по ЭЭГ
показателям мозг бодрствует, а организм спит. Отсюда и название парадоксального
сна.
Полагают, что парадоксальная фаза
сна — это аналог закрепившегося в эволюции состояния сторожевого бодрствования
и древнейшей формы активации, обнаруженной у холоднокровных в форме первичного
сна.
Особый интерес к стадии парадоксального
сна обусловлен возможностью изучать психофизиологические корреляты этого
состояния, которое увязывают с протеканием сновидений. Для классификаций стадий
сна у человека применяется схема Лумиса (рис. 3.36).
Для спокойного бодрствования
(стадия W) и дремоты характерен α—ритм, амплитуда которого постепенно
уменьшается и может вообще исчезать (стадия А). При внешнем раздражении
наступает реакция активации, т. е. происходит смена α—ритма на высокочастотный
(β—ритм (реакция десинхронизации). При появлении легкой сонливости (стадия
В) возникает неустойчивая высокоамплитудная активность. Стадию С обычно
связывают с поверхностным сном, когда начинают появляться
низкоамплитудные медленные волны. Переход к стадии D и особенно Е свидетельствует
о глубоком сне, который сопровождается дальнейшим усилением
высокоамплитудной медленной активности (δ—ритм).
|
|
|
Рис.
3.36 Классификация стадий сна у
человека по особенностям ЭЭГ Стадия W — бодрствование в расслабленном состоянии,
стадия А — переход от бодрствования ко сну, стадия В — засыпание и самый
поверхностный сон (острые вертекс—зубцы соответствуют «моменту» засыпания),
стадия С — поверхностный сон, стадия D — умеренно глубокий сон, стадия Е —
глубокий сон. Три нижние кривые представляют собой одновременную запись ЭЭГ,
электроокулограммы (ЭОГ) и электромиограммы указательного пальца (ЭМГ) во
время сна с быстрым движением глаз (БДГ) со сновидениями. |
МВС занимает у человека основную часть
сна — 75—85%, на долю ПС приходится 15—25%. Функции организма во время сна
существенно меняются. Их изменения во время МВС и ПС лучше рассматривать
раздельно, поскольку это разные состояния.
Функциональные характеристики
медленноволнового сна.
Системные изменения. Снижение: двигательной активности,
тонуса позных мышц, поведенческой реактивности, интенсивности метаболизма,
частоты сердечных сокращений и дыхания, вентиляции легких в ответ на СО2,
сосудистого тонуса, артериального давления, температуры «ядра» тела,
установочной точки терморегуляции, функции почек, моторики кишки.
Изменения в эндокринной системе. Уменьшение секреции кортизола и
тиреотропина, увеличение секреции гормона роста, альдостерона, тестостерона,
пролактина и инсулина, уровня гликогена в мозге.
Общие изменения нервной активности. Своей глубиной сон человека
значительно отличается от сна животных. Глубокие стадии МВС — чисто
человеческое приобретение, «артефакт цивилизации», что связано с безопасностью
ночлега и сохранностью от неожиданного нападения. Во время МВС могут возникать
приступы сомнамбулизма (от лат. somnus — сон и ambulo — бродить) (снохождения,
лунатизма), с этими же стадиями связаны ночные кошмары у детей и взрослых.
МВС, как правило, сопровождается сновидениями (см. разд. 3.23).
Изменения в головном мозгу. Замедление и рассредоточенность
мышления, короткие и редкие сновидения, снижение скорости метаболизма и
температуры мозга, региональные изменения кровотока и частоты разрядов
нейронов, гиперполяризация таламо—кортикальных нейронов, возникновение паттерна
разряда нейронов с паузой—вспышкой в некоторых отделах мозга, активизация
«сонно—активных» нейронов в переднем гипоталамусе, миндалевидном комплексе, в
ядрах основания переднего мозга и одиночного пути.
Во время сна многие процессы в
организме приостанавливаются; происходит снижение метаболизма, частоты
сердечных сокращений и дыхания, температуры мозга и тела. Однако экономия
расхода энергии во время сна в сравнении с бодрствованием в покое не велика —
всего 10—15%, что равняется одной не выпитой чашке молока для взрослого
человека.
Отчетливое снижение метаболизма
мозга происходит только во время δ—сна. Эти изменения никак не убеждают,
что сон предназначен для отдыха и экономии энергии. Увеличение секреции ряда
гормонов и уровня гликогена в мозге указывает на активацию анаболических
процессов в начале сна.
Функциональные характеристики
парадоксального сна.
Во время ПС, в отличие от МВС,
активность различных систем организма повышается: дыхание становится
нерегулярным, колеблется пульс и артериальное давление, повышается температура
мозга и скорость метаболизма. Еще один характерный признак ПС у мужчин —
появление эрекции, которая возникает даже у младенцев.
Сомато—висцеральные изменения. Быстрые движения глаз, сужение и
расширение зрачков, снижение чувствительности барорецепторов, вентиляции легких
в ответ на СО2, колебания частоты дыхания, пульса, артериального
давления, сосудистого тонуса, увеличение скорости метаболизма (у человека и
ряда видов животных), угнетение терморегуляторных ответов на тепло и холод,
активное расслабление скелетных мышц, эрекция.
Давно известно деление людей на два
типа — «утренний» («жаворонки») и «вечерний» («совы»). Одни легко просыпаются,
они энергичны с самого утра, для них утренние часы — наилучшее и самое
плодотворное время для работы. Люди «вечернего» типа с утра вялые,
раздражительные, им необходимо выпить кофе, чтобы ощутить прилив сил. Наивысшую
работоспособность они проявляют к вечеру, иногда к ночи. Полагают, что различия
между «жаворонками» и «совами» связаны с особенностями суточного ритма
температуры тела и висцеральных систем. В целом, сон «жаворонков» качественно
лучше, чем «сов». К тому же, в группе «жаворонков», как правило, доминируют
здоровые люди без вредных привычек.
Подавляющее большинство людей спит
от 7 до 9 ч в сутки. Только 5 человек из каждой тысячи спят 4—5 и менее часов.
К этой группе относятся Наполеон, Черчилль, Эдисон и некоторые другие известные
личности. А вот Эйнштейн, напротив, проводил в постели около 10 ч. Примерно 16
человек из тысячи спят более 10 ч. Очевидно, к этой группе относился и Илья
Ильич Обломов — герой одноименного романа И. А. Гончарова. Обследование 6 тыс.
близнецов, проведенное в Финляндии, показало, что на продолжительность сна
могут существенно влиять наследственные факторы.
Какая же длительность сна может
рассматриваться в качестве желательной «нормы» для здорового взрослого
человека? Эта «норма» достаточно широка, и часть сомнологов рекомендуют
индивидуальный режим сна. Однако исследование 1 млн человек в возрасте старше
30 лет заострило этот вопрос с самой неожиданной стороны. В группе людей,
спавших от 7 до 8 ч в сутки, был обнаружен наименьший уровень смертности, он
возрастал в 1,5—2 раза у долгоспящих (более 10 ч) и в 2,5 раза у малоспящих
(менее 4ч). И те, и другие
|
|
|
Рис. 3.37 Центры
сна (гипнотические центры) в стволе мозга кошки 1 — гипофиз, 2 — гипоталамус, 3 — мост, 4 — продолговатый мозг; штриховкой обозначены ядра, связанные со
сном без сновидений; зачерненный участок соответствует расположению нейронов,
контролирующих парадоксальный сон. |
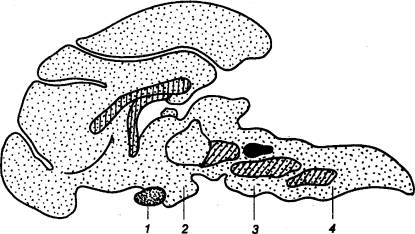
умирали от разных причин. Есть два
основных варианта интерпретации этих данных. Первый — слишком мало и слишком много
сна — одинаково плохо для здоровья и продолжительности жизни. Второй — очень
короткий и очень длинный сон лишь отражает различные состояния организма.
Наиболее глубокие изменения сна у
человека связаны с процессами созревания мозга и общего развития. Сон у
новорожденных определяется больше поведенческими, чем электрофизиологическими
критериями, потому что кора головного мозга младенца еще незрелая для
проявления явных признаков МВС и ПС. У маленьких детей эти состояния сна часто
называют «спокойным» сном (без движений глаз и тела) и «активированным». У
ребенка во время ПС постоянно наблюдаются подергивания рук, ног, мышц лица, а
недоношенные младенцы настолько беспокойны, что трудно разграничить состояния
ПС и бодрствования. Только к концу первого года жизни состояния сна можно
надежно определять при регистрации ЭЭГ.
ЭЭГ стадии МВС и ПС формируются в
онтогенезе в разные сроки, что может быть связано с неравномерностью развития
различных структур головного мозга: быстрее других достигают зрелости филогенетически
более древние образования (например, центры продолговатого мозга и моста) (рис.
3.37). Раньше других завершается формирование электрографической картины ПС,
несколько позднее формируется δ—сон. Позже всех отмечается становление ЭЭГ
стадии, характерной для начала сна.
Монофазный сон взрослых не является
обязательным, скорее всего он появился в процессе развития человеческой
цивилизации. Во—первых, в более жарких странах Средиземноморья, Центральной и
Южной Америки сохранился обычай спать днем в жаркое время суток («сиеста»).
Во—вторых, добровольцы, длительное время проживающие в глубоких пещерах, в
условиях изоляции от внешних факторов среды, постепенно переходят от
монофазного к полифазному типу сна.
Известно, что 60% пожилых людей в
возрасте от 65 до 83 лет спят один или даже два раза днем, часто пробуждаются и
бодрствуют среди ночи, т. е. возвращаются к полифазному типу сна, характерному
для младенцев.
Все без исключения млекопитающие
имеют сон и состояние МВС (табл. 3.3). ПС не обнаружен только у трех видов —
австралийской ехидны из отряда однопроходных яйцекладущих, афалины (большого
дельфина) и морской свиньи, или азовского дельфина. Для изучения сна
поведенческие критерии хотя и являются важными, но недостаточными. Состояние
неподвижности наблюдается у разных классов позвоночных и беспозвоночных
животных, а также у многих видов млекопитающих, впадающих в состояния
оцепенения и спячки; напротив, дельфины в течение суток двигаются, и у них нет
заметных периодов снижения активности. У многих видов животных идентифицированы
три состояния — бодрствование, МВС и ПС. Во время ПС наблюдаются ЭЭГ
десинхронизация в коре мозга, быстрые движения глаз, атония мышц шеи и
двуглавой мышцы бедра, а также гиперполяризация мотонейронов спинного мозга.
Как и у человека, сон млекопитающих
начинается с МВС и затем переходит в ПС. У мелких животных цикл сна (МВС—ПС)
короче, чем у крупных: у крысы он составляет 10 мин, у кошки — 30 мин, а у
слона — около 2 ч (судя по поведению). Для большинства животных характерен
многоразовый сон в течение суток.
Сон и зимняя спячка. Многие виды млекопитающих, для того
чтобы выжить в условиях дефицита пищи или воды, выработали гипометаболические
адаптации для периодического снижения расходов энергии — или в неактивную
фазу суток или в разные, неблагоприятные для их жизнедеятельности сезоны года.
Животные небольших размеров (бурозубки, мышовки и др.), а также крупные
(например, некоторые виды медведей) погружаются в разные формы оцепенения
(торпора) со снижением температуры тела до 30—25°С на периоды менее суток.
Суслики, сурки, летучие мыши и многие другие млекопитающие при недостатке пищи
впадают в более глубокие состояния зимней спячки (гибернации), снижая
температуру тела (мозга, сердца и других органов) часто почти до О °С на период
от 2 до 30 сут; при этом расход энергии сокращается у них в 10—100 и более раз.
Многие из них (например, суслики) в течение 6—8 зимних месяцев остаются без
пищи в глубоких норах в полной темноте и на холоде; 15—25 раз в течение зимы
они пробуждаются на 12—24 ч и вновь погружаются в спячку.
Зимой в таких условиях суслики,
например, имеют такое же количество МВС и ПС, как и летом в теплых условиях и
при достатке пищи. Очевидно, суровые зимние условия в результате адаптации
стали для них «комфортными», а МВС и ПС для них так же необходимы, как и летом.
В оцепенение и спячку они входят через МВС. В состоянии оцепенения и летней
спячки (эстивации), когда температура мозга поддерживается у сусликов на уровне
27—25 °С, ПС полностью исчезает, остаются только следы бодрствования и
регистрируется почти один МВС. Как известно, защитные терморегуляторные реакции
Таблица 3.3
Быстрый
сон у животных и человека
|
Вид |
Средняя
продолжительность одного цикла, мин |
Быстрый
сон, % от общей продолжительности сна |
|
Рептилии |
— |
Нет |
|
Курица,
голубь |
0,1 |
0—5 |
|
Сумчатая
крыса |
19 |
22—34 |
|
Крыса |
7—13 |
1—3 |
|
Кролик |
24 |
1—3 |
|
Овца |
— |
2—3 |
|
Кошка |
26 |
20—30 |
|
Обезьяна |
47—80 |
11—20 |
|
Человек |
75—85 |
15—25 |
во время ПС выключаются и поэтому его
сравнивают с состоянием короткой «пойкилотермии». В связи с этим
предполагается, что для сохранения строгой терморегуляции при оцепенении и
спячке ПС не нужен, даже вреден. В то же время в период МВС терморегуляторные
реакции сохранены, но значительно уменьшены, что и требуется для поддержания
температурного гомеостаза при более низком уровне температуры тела и
висцеральных функций.
Годовой максимум суточной
длительности ПС совпадает с периодом, предшествующим летней или зимней спячке,
когда животные поедают большие объемы пищи и резко увеличивают (в 1,5—2 раза)
массу тела и жировые резервы. Возможно, это увеличение времени ПС связано с
изменениями углеводного и липидного метаболизма в организме.
Наконец, максимальное увеличение
времени ПС обнаружено также перед каждым вхождением в спячку, в самом начале
снижения температуры мозга (до нижней границы «нормотермии», т. е. до 36—35
°С). При этом выявлены более длинные, ритмически повторяющиеся эпизоды ПС, во
время которых снижаются температура мозга, частота сердечных сокращений и
частота дыхания и увеличивается отток тепла через периферические сосуды.
Характерно, что локальное нагревание
предоптической области гипоталамуса, как одного из центров сна и
терморегуляции, моделирует эти изменения ПС и также способствует снижению
температуры мозга. Снижение температуры мозга во время ПС выявлено только у
видов, впадающих в оцепенение и спячку. У «нормотермных» животных — кошек,
крыс, собак, мышей, кроликов — во время ПС она, напротив, повышается. Для этой
большой группы животных общим являются следующие биологические признаки: 1)
незрелость при рождении; 2) более низкая скорость основного метаболизма, низкий
среднегодовой уровень энерготрат и температуры тела; 3) безопасные условия сна
(глубокие норы и другие надежные укрытия); 4) большое количество сна.
Изучение экологической группы водных
и полуводных млекопитающих, включающей представителей отрядов китообразных,
сирен и ластоногих, привело к поразительным открытиям, демонстрирующим влияние
изменившихся в ходе эволюции экологических условий обитания на структуру сна.
Сон морских млекопитающих существенно отличается от сна наземных, причем у
разных экологических групп в процессе эволюции были использованы разные пути
адаптации ко сну в воде. Максимальные отличия выявлены у дельфинов: 1) сон не
сопровождается обязательной неподвижностью и может проявляться на фоне
постоянной двигательной активности при плавании; 2) доминирующей формой сна
является однополушарный МВС, на который приходится 29% времени суток; глубокий МВС
(δ—сон) никогда не возникает в обоих полушариях одновременно; 3)
классический ПС не наблюдается, левая и правая половины мозга спят по очереди.
У разных видов ластоногих в отличие
от китообразных, выявлены два основных типа сна — «наземный» (преобладание
одновременного развития МВС в обоих полушариях и наличие классической картины
ПС) и «водный» (межполушарная асимметрия МВС, приуроченность сна к дыхательным
паузам). Это обусловлено тем, что эти животные являются полуводными и
адаптированы к обитанию и сну в двух средах. Северные морские котики большую
часть времени в лежбищный период (июнь—сентябрь) проводят на суше, а осенью
уходят в открытое море и вынуждены спать только в воде. Отличительной чертой
сна в воде у них является выраженная межполушарная асимметрия протекания МВС,
что связано с постоянной двигательной активностью только одной передней ласты,
поскольку они лежат на боку и ноздри у них все время находятся над поверхностью
воды.
Таким образом, у водных и полуводных
млекопитающих выявлены три разных типа сна, что демонстрирует пластичность
механизмов сна в процессе эволюции и его экологическую детерминированность.
Общая особенность морских обитателей — малое количество сна. Если из суточного
МВС исключить состояние дремоты, то каспийские тюлени спят меньше осла и
лошади, т. е. они являются «рекордсменами» по минимальному количеству сна среди
всех млекопитающих.
Сон у птиц. Наиболее часто наблюдаемые позы сна
у птиц — голова опущена на грудь или повернута назад, клюв лежит на крыле или
под ним, животное стоит на двух или одной ноге. Птицы, утерявшие способность
летать (например, страус), часто спят на земле, вытянув шею и ноги вдоль
туловища. Плавающие птицы могут спать на плаву. Предполагается, что некоторые
птицы могут спать во время парения или длительных перелетов, однако такой
феномен пока не подтвержден электрографическими показателями. Периодическое
открывание глаз во время сна обусловлено фактором опасности. Сон в стае
(«коллективный» сон), по—видимому, более спокойный и более продолжительный.
Обыкновенная пустельга спит с одним открытым глазом, а сова вообще не закрывает
глаз. Большинство птиц, исключая таких ночных хищников, как сова, спят ночью
(голубь, скворец, ястреб). Продолжительность сна у разных видов колеблется от 1
до 16 ч в сутки. У всех птиц, как и у млекопитающих, для МВС характерны
медленные волны, а для ПС — ЭЭГ десинхронизация и быстрые движения глаз. У ряда
птиц (голубь, черный дрозд) также отмечено максимальное общее время МВС в
начале ночи, а ПС — во второй ее половине.
Эволюция сна. Различные виды черепах,
крокодилов и ящериц имеют характерные паттерны поведения,
соответствующие общепринятым критериям поведенческого сна: прекращение
двигательной активности, принятие соответствующей виду позы покоя, расслабление
мышц, повышение порогов на внешние стимулы, быстрое пробуждение и возвращение к
поведению, типичному для состояния бодрствования. Однако в исследованиях с
регистрацией электрической активности различных структур мозга рисунки ЭЭГ
существенно отличаются от. ЭЭГ во время сна у высших позвоночных. Наиболее
часто наблюдаются или незначительные изменения в ЭЭГ при переходе к
поведенческому покою, сходному со сном, или высокоамплитудные «всплески»
ЭЭГ—волн; последние, возможно, являющиеся аналогами ЭЭГ—волн, возникают в вентральном
гиппокампе во время МВС у некоторых млекопитающих. Значительные отличия этих
«всплесков» ЭЭГ—волн от картины МВС у птиц и млекопитающих могут быть
обусловлены отсутствием у рептилий новой коры, в которой проявляются медленные
волны.
Земноводные (лягушки, жабы и саламандры)
длительное время остаются неподвижными во время бодрствования, и поэтому
определить у них состояния покоя, сходные со сном, еще труднее, чем у рептилий.
В тех случаях, когда это удается сделать, небольшие изменения частоты и амплитуды
ЭЭГ никогда не обнаруживают какого—либо заметного сходства с
электрографическими показателями сна у высших позвоночных.
Рыбы, характеризующиеся более широким
видовым разнообразием, совершенно неожиданно оказались более удачными объектами
для выявления признаков поведенческого сна. Некоторые из них подыскивают
подходящие места для своих «водных постелей» и принимают характерную для сна
позу: зарываются в ил, отдыхают на скалах, заплывают в пещеры в скалах или
неподвижно висят в воде в разных позах, в том числе на боку или даже вверх
брюхом. В таком состоянии рыбы обычно реагируют только на сильные внешние
стимулы. Рыба—чистильщик, отдыхающая в коралловых нишах, и рыба—попугай
выделяют много слизи и прячутся в такой капсуле, защищающей их от врагов во время
отдыха. В большинстве исследований не обнаружено корреляции ЭЭГ с такими
состояниями покоя у рыб.
У насекомых (пчел, бабочек),
у разных видов скорпионов и моллюсков также наблюдаются состояния
покоя, сопровождающиеся расслаблением мышц и повышением порогов реакции на
внешние стимулы. Состояния поведенческого покоя, характерные для всех
пойкилотермных организмов (низших позвоночных и беспозвоночных), существенно
отличаются от состояний истинного сна, накладывающихся на состояния покоя у
гомойотермных животных.
Сон имеет сложную нейрогуморальную
природу. В истории изучения сна наибольший интерес представляют гуморальные
и нервные теории сна. Еще в начале XX в. было высказано предположение о
том, что сон возникает в результате накопления токсических продуктов обмена,
накапливающихся вследствие дневного утомления (гипнотоксины). Оказалось
также, что экстракты мозга таких животных и находящихся в зимней спячке
вызывают у кошек и собак длительное сонное состояние. Если, используя препарат
с перекрестным кровообращением двух кроликов, у одного раздражать участки
мозга, вызывающие сон, то второй тоже засыпает. Это связывалось с переносом
каких—то гипногенных веществ, в избытке накапливающихся в организме
(ацетилхолин, ГАМК, серотонин).
Уже с середины XIX в. устойчивые
позиции занимают нервные теории сна. Клиника уже давно накапливала
материал о связи гипоталамуса с регулированием процессов сна. Описано даже
существование в гипоталамусе так называемого центра сна. И. П. Павлов не
соглашался со взглядами о подкорковой природе сна, отдавая предпочтение
корковым механизмам. И. П. Павлов рассматривал сон как следствие иррадиации
торможения, распространяющегося по коре больших полушарий, и находил много
общего между активным условнорефлекторным сном и условным торможением.
Существует ли в мозге какой—либо
«центр», ответственный за наступление и окончание бодрствования и сна? После
Первой мировой войны в Европе много жизней унес эпидемический летаргический
энцефалит, одним из симптомов которого был длительный летаргический сон. В этом
случае австрийский невропатолог К. Экономо описал воспалительные изменения
вокруг водопровода среднего мозга и в задней стенке III желудочка мозга, а
также выдвинул теорию о «центре» сна. С учетом работ И. П. Павлова он полагал,
что сон возникает в результате разлитого торможения, распространяющегося на
таламус, кору мозга и другие структуры. После работ, открывших активирующие
влияния ретикулярной формации ствола мозга, были проведены многочисленные
исследования, которые выявили участие многих подкорковых структур в регуляции
бодрствования и сна. Но ни одна из структур, согласно современным
представлениям, не может претендовать на роль центра, регулирующего эти
состояния.
Сегодня уже с несомненностью установлена
корково—подкорковая локализация структур управления сном (рис. 3.37, 3.38).
Разработана гипотеза о моноаминергической регуляции стадий сна.
Медленноволновый сон регулируется серотонинергической системой ядер шва в
продолговатом мозгу: установлена прямая зависимость сна от функциональной
активности этих мозговых образований, общего уровня серотонина и его обмена. Катехоламинергические
нейроны голубоватого пятна и ретикулярной формации среднего мозга
выступают как антагонистическая система, ответственная за возникновение
быстрого сна и состояния бодрствования.
Предполагается, что в системы
поддержания состояния бодрствования включены тормозные нейроны разных отделов
мозга. Их активность возрастает при ослаблении функции активирующих нейронов и
способствует дальнейшему
|
|
|
Рис. 3.38 Представление о нейрофизиологии и нейрохимии сна А — бодрствование; Б — медленноволновая фаза сна; В —
парадоксальный сон. |
ослаблению деятельности последних. Активирующие
нейроны, поддерживающие состояние бодрствования, выделяют множество
передатчиков: глутаминовую кислоту, ацетилхолин, норадреналин, серотонин,
гистамин. В отличие от этого, тормозные нейроны, по—видимому, секретируют лишь
один основной химический агент — ГАМК, которая рассматривается как главный
тормозный медиатор мозга. Следовательно, «центра» МВС нет, а наступление
дремоты и МВС зависит от периодического ослабления деятельности нейронов
подкорковых образований, оказывающих тонические активирующие влияния на кору
мозга.
Существуют указания на особую роль
глиальных элементов во время сна. Одной из основных нейрохимических функций
медленного сна являются пластические репарационные процессы, связанные с
метаболизмом белков и РНК, причем главным образом в глиальных клетках
определенных структур мозга. Активные конформационные изменения в мембранах
нейронов при парадоксальном сне являются одной из причин своеобразия ЭЭГ при
данном функциональном состоянии.
Таким образом, в эволюции четко
обозначены три различные формы деятельности мозга: бодрствование, быстрый сон и
медленный сон.
Нарушения сна и бодрствования. Искусственное лишение человека сна
является тяжелейшим испытанием. У животных, которые погибали в результате
лишения сна, обнаруживали кровоизлияния в коре больших полушарий, в стволе и
спинном мозгу.
Нарушения сна особенно
распространены среди населения цивилизованных стран. У некоторых людей ощущение
отсутствия сна может возникать даже тогда, когда они находятся в глубокой
стадии сна. Американские ученые исследовали людей с якобы плохим сном для
выявления причин таких ложных ощущений. Оказалось, что у этих людей нарушается
ночью реальный отсчет времени, или во сне протекает активная психическая
деятельность, или, наконец, имеют место качественные, а не количественные
нарушения сна. Последнее зависит от дефицита глубокого медленного сна и частых
ночных пробуждений. Следовательно, у части лиц действительно есть объективные
основания быть неудовлетворенными своим сном. Основными причинами бессонницы
считают недостаток мышечной деятельности, преклонный возраст и нарушения
традиционного суточного ритма, вызванного цивилизацией.
Главную роль в нарушениях сна играют
неврозы, в борьбе с которыми первое место занимает психотерапия, затем
физиотерапия (электросон), наконец, химиотерапия (психофармакология),
заключающаяся в недифференцированном использовании снотворных веществ, которые
подавляют быстрый сон, сопровождающийся сновидениями.
3.14.3. Закономерности
условнорефлекторной деятельности
Характерная особенность условных
рефлексов состоит в том, что они образуются в течение индивидуальной жизни
организма и сохраняются постоянно. Они могут исчезать и вновь появляться в
зависимости от изменяющихся условий среды и состояния организма. Временный
характер условного рефлекса обеспечивается наличием процесса торможения,
который наряду с процессом возбуждения определяет динамику условнорефлекторной
деятельности.
Общие признаки условных рефлексов. 1. Условные рефлексы носят приспособительный
характер, что делает поведение наиболее пластичным, подогнанным к
конкретным условиям среды. 2. Любые условные рефлексы требуют для своего
участия высших отделов головного мозга (у насекомых, например, головных
ганглиев — грибовидных тел). 3. Условные рефлексы приобретаются и отменяются в индивидуальной
жизни каждой конкретной особи. В индивидуальную память записываются не
всякие случайные совпадения условного раздражения и безусловной реакции, а лишь
те, сочетание которых во времени оказывается наиболее вероятным. Все остальные
случайно возникшие связи затормаживаются. 4. Условный рефлекс имеет сигнальный
характер, т. е. предшествует, предупреждает последующее возникновение
безусловного рефлекса, подготавливая к нему организм. С помощью условных
рефлексов животное может заранее избежать опасности или подготовиться к захвату
добычи, встрече с половым партнером и т. д.
Таким образом, условные рефлексы
— это индивидуально приобретенные системные приспособительные реакции
животных и человека, возникающие на основе образования в центральной нервной
системе временной связи между условным (сигнальным) раздражителем и
безусловнорефлекторным актом.
Классификация условных рефлексов. Существует несколько систем
классификации условных рефлексов, каждая из которых строится на одном ведущем факторе,
положенном в ее основу.
I. Условные рефлексы различаются по особенностям
безусловного подкрепления. В зависимости от наличия или отсутствия
подкрепления условные рефлексы делят на положительные (подкрепляемые),
вызывающие соответствующую реакцию организма, и отрицательные, или тормозные
(неподкрепляемые), которые не только не вызывают соответствующей реакции, но и
ослабляют ее.
Согласно биологическому значению
подкрепления, условные рефлексы разделяют в зависимости от биологических
потребностей. Различают витальные условные рефлексы (пищевые,
оборонительные, регуляции сна и пр.), зоосоциальные (половой,
родительский, территориальный и пр.) и условные рефлексы саморазвития
(исследовательский, имитационный, игровой и др.).
Иногда имитационные (подражательные)
условные рефлексы, подкреплением в которых служит лишь наблюдение за поведением
другой особи, выделяют в самостоятельную группу.
Подкреплением условному рефлексу
может служить любая деятельность организма. Достаточно грубо ее можно разделить
на две большие группы: двигательные и автономные (вегетативные) условные
рефлексы. На базе двигательных реакций образуется множество
инструментальных условных рефлексов, которые в одних случаях могут копировать
безусловный рефлекс, в других — иметь гораздо более сложную форму и служить
основой произвольного поведения.
К вегетативным условным
рефлексам относится классический слюноотделительный условный рефлекс.
Условнорефлекторному контролю могут подчиняться почти все внутренние органы.
Целостный условный рефлекс всегда
имеет полиэффекторную природу, и отнесение его к той или иной категории
рефлексов зависит от того, какой из эффекторов регистрируется
экспериментатором.
Различаются условные рефлексы и по характеру
подкрепления. Если в качестве подкрепления используется простой безусловный
рефлекс, то вырабатываются условные рефлексы первого порядка. Если же
подкреплением служит ранее выработанный прочный условный рефлекс, то
образующуюся связь именуют условным рефлексом второго порядка. Соответственно
возможны условные рефлексы более высоких порядков. Именно они легко
вырабатываются у человека и составляют основу развития мыслительной
деятельности.
II. По характеру условного
сигнала условные рефлексы подразделяют на две большие группы: экстероцептивные
и интероцептивные. Среди экстероцептивных выделяют зрительные, слуховые,
обонятельные, вкусовые, тактильные, температурные условные рефлексы в
соответствии с рецепторным прибором, на который действует раздражитель.
Обучение речевому общению между людьми в основе своей имеет специально
человеческие условные рефлексы на словесные сигналы.
Интероцептивные условные рефлексы возникают в том случае, если
поступивший в головной мозг афферентный залп из внутренних органов становится
сигналом к вегетативной или двигательной деятельности организма. Условные
рефлексы от внутренних органов разделяют на механические, химические,
температурные, осмотические и др.
По природе условного сигнала
различают натуральные и искусственные условные рефлексы. К натуральным
условным рефлексам относят такие, которые образуются на естественные
признаки безусловного раздражителя (например, вид и запах мяса). Любые условные
сигналы, не являющиеся, например, непременным свойством пищи, а лишь
совпадающие с моментом ее поедания, относятся к искусственным условным
сигналам.
По структуре условного сигнала
различают условные рефлексы: а) на простые мономодальные раздражители; б) на
одновременные комплексные раздражители, состоящие из нескольких компонентов
(например, свет + звук + кожное раздражение); в) на последовательные комплексы;
г) на цепи раздражении, компоненты которых действуют последовательно, не
совпадая друг с другом, а безусловное подкрепление присоединяется лишь к
последнему из них.
Иногда в качестве условных сигналов
используют не абсолютные, а относительные признаки раздражителей.
Например, из двух одновременно предъявляемых однородных раздражителей — плоских
кружков — условный рефлекс вырабатывается на один из них — больший по размеру.
Затем животному предъявляют геометрические фигуры (например, треугольники),
также отличающиеся по размеру. Если и в этом случае животное осуществит
условнорефлекторную реакцию на сигнал более крупного размера, судят о выработке
у него условного рефлекса на отношение.
III. Важнейшим признаком
классификации является соотношение во времени действия условного и
безусловного раздражителей. По этому признаку различают: а) наличные
условные рефлексы, образующиеся при совпадении времени действия сигнала и
подкрепления, и б) следовые условные рефлексы, при образовании которых сигнал и
подкрепление отделены временным интервалом.
Если подкрепление почти сразу
присоединяется к сигналу, образуются совпадающие условные рефлексы, а
если спустя 5—30 с от начала его действия — вырабатываются отставленные
условные рефлексы. Условные рефлексы, при которых подкрепление присоединяется к
сигналу после длительного изолированного действия последнего, называются запаздывающими
условными рефлексами.
Особая разновидность следовых
условных рефлексов — это условные рефлексы на время. Если какой—либо
безусловный рефлекс проявляется через одинаковые интервалы времени, то в
результате каждый раз по истечении данного интервала как бы сама по себе
возникает реакция, вызывавшаяся ранее действием безусловного раздражителя. Такая
реакция получила название условного рефлекса на чистое время.
Стадии образования условных
рефлексов.
Образование условного рефлекса (рис. 3.39) обычно проходит две стадии:
генерализации и специализации. На первой стадии— генерализации —
условнорефлекторное действие приобретают не только подкрепляемый условный
сигнал, но и сходные с ним раздражители. Например, у собаки вырабатывается
пищевой условный рефлекс на тон 400 Гц. Побежка животного к кормушке по этому
сигналу подкрепляется пищей. Но близкие сигналы, например тон 200 Гц и тон 600
Гц, также вызывают побежку к кормушке, хотя и не сопровождаются пищевым
подкреплением. Весьма широкий набор близких к подкрепляемому раздражителей
приобретают на первой стадии выработки условного рефлекса значение условных
сигналов. Происходит как бы первичная сенсорная генерализация, обобщение
признаков условных сигналов.
Степень генерализации зависит от
многих факторов и прежде всего от модальности и характера условного сигнала,
безусловного подкрепления и их временного сочетания. Биологическое значение
стадии генерализации чрезвычайно велико, ибо в естественных условиях обитания
позволяет животному адаптироваться сразу к широкому спектру сходных сигналов.
И. П. Павлов эту ситуацию
образования условных рефлексов объяснял широкой иррадиацией возбудительных
процессов в пределах анализатора, воспринимающего условный сигнал. А. А.
Ухтомский полагал, что на первой стадии образования условный рефлекс протекает
по механизмам доминанты, когда неподкрепляемые посторонние раздражители
способны активизировать поведение животного по удовлетворению доминирующей
биологической потребности.
Вторая стадия заключается в дифференцировке
условнорефлекторного ответа лишь на подкрепляемый сигнал, тогда как все
остальные сходные по качеству раздражители становятся неэффективными.
|
|
|
Рис. 3.39 Схемы образования условного рефлекса по И. П. Павлову (А) и Э. А.
Асратяну (Б) К — кора; ЗЦ — зрительный центр; СЖ — слюнная железа; КП —
корковое представительство пищевого центра; Г — глаз; М — мышца; Я — язык; Ц
— подкорковый центр: 1 — временная связь между зрительным и
слюноотделительным рефлексами через серое вещество коры большого мозга; 2 —
через ассоциативные пути белого вещества; 3 — через подкорковые центры. |
Скорость наступления этой стадии и
степень специализации, как и в предыдущей фазе, зависят от всех трех факторов:
сенсорного и эфферентного звеньев условного рефлекса, а также временных соотношений
их активации. Стадия специализации условного рефлекса обычно совпадает с
началом его автоматизации.
Биологически эта стадия способствует
выделению из широкого репертуара возможных только узкого набора значимых
сигналов, имеющих наибольшую вероятность запуска безусловнорефлекторной
реакции. И. П. Павлов эту стадию объяснял процессами концентрации возбуждения в
анализаторе условного сигнала. Эта гипотеза пока не получила подтверждения
современными нейрофизиологическими методами. Снижение эффективности
неподкрепляемых раздражителей И. П. Павлов объяснял также выработкой
внутреннего условного торможения, нейрофизиологические механизмы которого еще
ждут своей окончательной разработки.
3.14.4. Торможение условных
рефлексов
И. П. Павлов, изучая условные
рефлексы и их взаимоотношения, наблюдал торможение (угнетение) условных
рефлексов при действии посторонних или сильных раздражителей, а также слабых —
при болезненном состоянии организма. Он считал, что баланс между возбуждением и
торможением определяет внешнее проявление поведения животных и человека, и
выдвинул собственную схему классификации видов торможения при
условнорефлекторной деятельности.
Внешнее (безусловное) торможение. Под внешним торможением
понимают срочное подавление текущей условнорефлекторной деятельности при
действии посторонних для нее раздражении, вызывающих ориентировочный или
какой—либо другой безусловный рефлекс. По механизму своего возникновения этот
тип торможения относят к врожденным, которые осуществляются благодаря
явлениям отрицательной индукции (индукционное торможение, по Павлову).
А. А. Ухтомский именовал его сопряженным торможением и видел в нем
физиологическую основу для выполнения доминирующей формы деятельности
организма. Безусловное торможение называют внешним потому, что причина его
возникновения лежит вне структуры самого тормозного рефлекса.
Ориентировочный рефлекс — наиболее часто встречающийся фактор
безусловного торможения. Однако тормозный эффект ориентировочного рефлекса при
повторении того же сигнала постепенно
ослабевает и может исчезнуть полностью. При этом и сам ориентировочный рефлекс
перестает наблюдаться. Ориентировочный рефлекс («что такое?») возникает
для более полного восприятия информации, содержащейся в неожиданном и
постороннем раздражителе.
В обиходе постоянно наблюдается, как
человек прекращает текущую деятельность в результате переключения внимания на
новый внезапно появившийся раздражитель. В момент возникновения этого рефлекса
проявляется сопряженное торможение конкурирующих рефлексов. Оно может быть
более или менее глубоким, кратковременным или более длительным, что зависит от
физиологической силы ориентировочного и тормозного рефлексов. При повторном
раздражении в силу привыкания ориентировочный рефлекс пропадает, одновременно
снижается и эффект внешнего торможения. Такой вид торможения был назван гаснущим
тормозом.
Другой вид безусловного торможения
отличается постоянством своего эффекта на тот или иной тормозимый рефлекс и
поэтому называется постоянным тормозом. Стабильность внешнего торможения
определяется, в частности, физиологической силой тормозящего рефлекторного
акта. К жизненно важным для организма рефлексам относят оборонительные
безусловные рефлексы на разные вредящие раздражения, включая болевые. Как и в
случае гаснущего тормоза, длительность постоянного тормоза оборонительного
рефлекса определяется его силой и характером тормозимого рефлекса и, в
частности, степенью его упроченности.
«Молодые» условные рефлексы
тормозятся легче и на более длительный срок, чем более «старые», при одних и
тех же условиях. Нетвердо заученные поведенческие навыки или знания легче
исчезают при сильном неприятном постороннем воздействии, чем более твердо
усвоенные жизненные стереотипы. Болевые воздействия от внутренних органов
обладают более длительным тормозным влиянием на условнорефлекторную
деятельность. А иногда их сила настолько велика, что извращает нормальное
протекание даже безусловных рефлексов.
Следовательно, два антагонистических
рефлекса — пищевой и оборонительный — не могут сосуществовать, более слабый
тормозится под влиянием более сильного.
В этом плане павловское внешнее
торможение выступает в качестве тончайшего инструмента, способного выделить
наиболее биологически значимую форму поведения, подчинив ей все остальные виды
деятельности. С позиций учения о доминанте это можно рассматривать как
сопряженное торможение при доминанте, которое выполняет определяющую роль при
ее формировании. И это торможение должно быть своевременным, т. е. иметь
координирующее значение для работы других органов и организма в целом.
Хорошо известно, что если
увеличивать интенсивность какого—либо раздражения, то вызываемый им эффект
увеличивается (закон силы}. Однако дальнейшее усиление раздражения
приведет к падению или полному исчезновению эффекта. В основе этого результата
лежит не утомление, а запредельное торможение, которое И. П. Павлов
назвал охранительным, так как оно ограждает клетки мозга от избыточного
расходования энергетических ресурсов. Этот вид торможения зависит от
функционального состояния нервной системы, возраста, типологических
особенностей, состояния гормональной сферы и пр.
Предел выносливости клетки в
отношении раздражителей разной интенсивности называется пределом ее
работоспособности, и чем выше этот предел, тем легче клетка переносит
действие сверхсильных раздражителей. Причем речь идет не только о физической,
но и об информационной силе (значимости) условных сигналов.
Крайним случаем запредельного
торможения является оцепенение, возникающее у животных и человека под влиянием
сверхсильного раздражения. Человек может впасть в состояние ступора —
обездвиженности с полным или частичным отсутствием речевого общения. Такие
состояния возникают не только в результате действия физически сильного
раздражителя (взрыва бомбы, например), но и вследствие тяжелых моральных
потрясений (например, при неожиданном сообщении о тяжелой болезни или смерти
близкого человека — психогенный ступор).
Внутреннее (условное) торможение. К форме внутреннего торможения условнорефлекторной
деятельности относят те ее случаи, когда условный раздражитель перестает
подкрепляться безусловным, т. е. постепенно теряет свое пусковое сигнальное
значение. Такое торможение возникает не срочно, не сразу, а развивается
медленно по общим законам условного рефлекса и является столь же изменчивым и динамичным.
И. П. Павлов назвал его поэтому условным торможением. Он считал, что
такое выработанное торможение возникает внутри центральных нервных структур
самих условных рефлексов, а отсюда и его название — внутреннее (т. е. не
наведенное извне, не индукционное).
Выделим основные черты
условного торможения. Оно: 1) развивается при неподкреплении раздражителей,
которые постепенно приобретают свойства условного тормозного или отрицательного
сигнала. 2) поддается тренировке. Заторможенный условный рефлекс может
самопроизвольно восстанавливаться, и это свойство чрезвычайно важно при
воспитании поведенческих навыков в раннем возрасте. 3) зависит от
физиологической силы безусловного рефлекса, подкрепляющего положительный
условный сигнал. 4) зависит от прочности ранее выработанного условного
рефлекса. 5) может взаимодействовать с безусловным, в этих случаях возникает
явление растормаживания, а иногда в результате суммации условного и
безусловного торможений их общий эффект может усиливаться 6) способность к
разным проявлениям условного торможения зависит от индивидуальных свойств
нервной системы: у возбудимых индивидуумов оно вырабатывается труднее и
медленнее. И. П. Павлов подразделил условное торможение на четыре вида:
угасательное, дифференцировочное, условный тормоз, торможение запаздывания.
Угасательное торможение развивается при отсутствии
подкрепления условного сигнала безусловным. Условные рефлексы обладают
временным характером именно потому, что при отмене безусловного подкрепления
соответствующая мозговая связь теряет свою силу, тормозится иногда надолго, а
иногда и вовсе перестает существовать.
Представим, что вид определенной
местности постоянно сочетается у животного с получением пищи. Но если здесь пищевые
ресурсы исчезли, животное со временем, не найдя пищи, перестанет посещать ранее
знакомую местность вследствие развития угасательного торможения. Величина и
скорость выработки угасательного торможения зависят от прочности условного
рефлекса (стабильные рефлексы угашаются медленнее), от физиологической силы и
вида безусловного рефлекса (угащение у голодной собаки труднее, чем у сытой;
пищевые условные рефлексы угашаются
быстрее, чем оборонительные), от частоты неподкрепления (регулярное
неподкрепление способствует быстрому развитию торможения). Оно развивается
волнообразно и зависит от индивидуальных типологических различий.
Дифференцировочное торможение развивается при неподкреплении
раздражителей, близких по свойствам к подкрепляемому сигналу. Этот вид
торможения лежит в основе различения раздражителей. С помощью
дифференцировочного торможения из массы сходных раздражителей выделяется
биологически для него важный, а на другие похожие раздражители условная реакция
будет выражена слабее или отсутствовать полностью.
Свойство генерализации
(первичного обобщения) условных рефлексов — неизбежный атрибут поведенческих
адаптации животных в естественной среде обитания. Учитывая, что изменчивость
окружающей среды происходит по вероятностному закону и колебания тех или иных
биологически значимых признаков предвидеть с высокой вероятностью невозможно,
становится биологически оправданной значительная сенсорная генерализация
условных рефлексов на стадии активного поиска жизненно важных объектов.
На стадии генерализации условных
рефлексов обнаруживается механизм доминанты., одной из характерных черт
которой является способность рефлекторной системы диффузно отзываться на
широкий репертуар внешних стимулов. В процессе повторного осуществления данного
рефлекторного акта диффузная отзывчивость сменяется избирательным реагированием
только на те раздражения, которые первично создали данную доминанту. Стадия
специализации доминанты наступает благодаря механизмам дифференцировочного
торможения.
Последнее обладает следующими основными
свойствами: 1) чем ближе дифференцируемые раздражители, тем труднее выработать
на один из них дифференцировочное торможение; 2) степень торможения
определяется силой возбуждения, развиваемого положительным условным рефлексом;
3) выработка этого торможения происходит волнообразно; 4) дифференцировочное
торможение тренируемо, что лежит в основе тонкого распознавания сенсорных
факторов среды.
В самостоятельный вид условного
торможения И. П. Павлов выделил условный тормоз, который образуется при
неподкреплении комбинации из положительного условного сигнала и индифферентного
раздражителя. Например, у собаки образован пищевой условный рефлекс на звук.
Если к этому сигналу присоединить свет лампочки и их совместное действие не
подкреплять пищей, то после нескольких применений эта комбинация перестанет
вызывать пищевую реакцию, хотя изолированное применение звонка по—прежнему
будет вызывать обильное слюноотделение. По существу, это вариант
дифференцировочного торможения.
Прибавочный раздражитель в первый
момент своего применения в комбинации с положительным сигналом вызывает
ориентировочный рефлекс и торможение условной реакции (внешнее торможение),
затем превращается в индифферентный раздражитель (гаснущий тормоз), и, наконец,
на месте безусловного торможения развивается условный тормоз. Если прибавочный
раздражитель приобрел эти свойства, то, будучи присоединенным к любому другому
положительному сигналу, он затормозит соответствующий этому сигналу условный
рефлекс.
При выработке торможения
запаздывания подкрепление соответствующим безусловным рефлексом не
отменяется, как в предыдущих видах торможения, а значительно отодвигается от
начала действия условного раздражителя. Подкрепляется лишь последний период
действия условного сигнала, а предшествующий ему значительный период его
действия лишается подкрепления. Именно этот период сопровождается торможением
запаздывания и называется недеятельной фазой запаздывающего условного
рефлекса. По его истечении торможение прекращается и сменяется возбуждением
— так называемая деятельная фаза рефлекса. В данном случае участвуют два
раздражителя в комплексе и второй компонент — время.
В экспериментах с пищевыми условными
рефлексами отставление подкрепления от начала условного сигнала может достигать
2—3 мин, а при электрооборонительных — 30—60 с. Адаптивное значение торможения
запаздывания состоит в тонком анализе времени отставления раздражителя,
положительная фаза рефлекса приурочивается ко времени запуска безусловного
рефлекса. Например, кошка, поджидающая жертву у мышиной норки, не обнаруживает
слюноотделения до тех пор, пока мышь не окажется у нее в зубах.
Тесное взаимодействие разных видов
условного торможения, в особенности условного и безусловного торможений, а
также возможность выработки условного торможения на базе безусловного являются
убедительным основанием для предположения об их единой физиологической природе.
3.15. ОСНОВНЫЕ МЕХАНИЗМЫ РАБОТЫ
МОЗГА
3.15.1. Механизмы формирования
условных рефлексов. Теория конвергенции
Ч. Шеррингтон сформулировал
фундаментальный закон о работе нервной системы — принцип общего пути,
согласно которому эфферентный нейрон является общим путем для импульсов,
возникающих в любом из многочисленных источников рецепции. Подсобная
координация складывается на основе морфологического преобладания афферентных
путей над эфферентными. Так формируется и реализуется механизм конвергенции
— схождения множества афферентных путей в единственный анатомически
ограниченный эфферентный канал.
А. А. Ухтомский распространил этот
принцип конвергенции на все этажи ЦНС, полагая, что в онтогенезе и филогенезе
конвергенция все более расширяется и обогащается по содержанию, возрастает
скорость интерференции рефлекторных действий в общем пути. И если у Шеррингтона
рассмотрен принцип статической, морфологической конвергенции путей, то
Ухтомский в последующем при разработке учения о доминанте обосновал принцип
подвижной, физиологической конвергенции нервных влияний по отношению к области
преобладающего текущего действия. Именно это и послужило основой его учения
о доминанте как важнейшем принципе интегративной деятельности мозга.
А. А. Ухтомский отмечал тесную связь
между учением Ч. Шеррингтона о конвергенции и И. П. Павлова об условных
рефлексах, которому удалось показать само закладывание и образование связей в
«воронке», привлечение все новых афферентаций, конвергирующих к эфферентным
(исполнительным) приборам.
Конвергенция гетеросенсорных
стимулов была положена в основу понимания механизмов условных рефлексов. Дж.
Эклс (1953) предположил существование нелокализованного центра конвергенции,
где сходятся условный и безусловный раздражители. Их взаимодействие и
интеграция завершаются формированием особой пространственно—временной
структуры, импульсных разрядов. Многократные сочетания включают механизм
потенциации путей условного сигнала который приобретает теперь способность
запускать условнорефлекторный акт. Хранение этого эффекта обеспечивается
механизмами реверберации возбуждения в нервных сетях. Д. Албе—Фессар
(1964) полагал, что конвергенция, фоновая ритмическая активность и потенциация
образуют некоторую цепь динамических процессов, связывающих интеграцию в
пространстве (обусловленную конвергенцией) с интеграцией во времени
(обусловленную потенциацией).
Решающим фактором в механизмах
образования условных рефлексов выступает биологическое значение
подкрепляющего рефлекса. Свойства подкрепляющего фактора определяются
специфическими химическими реакциями, которые он вызывает, и соответствующими
молекулярными преобразованиями в пределах мембраны и цитоплазмы нейрона. Здесь
в результате ультраструктурных и молекулярных преобразований и происходит
«сцепление» условной и безусловной стимуляции с последующим выходом сложившейся
интеграции на аксон в виде клеточного разряда (рис. 3.40).
На рис. 3.41 показано, что синаптические входы для условного и
безусловного стимулов конвергируют на пресинаптической («входной») клетке в
цепи нейронов. Сложившаяся ассоциация («интеграция») передается на
постсинаптический нейрон («выходной»), но по одному общему пути и для
сигнального, и для подкрепляющего раздражителей.
|
|
||
|
Рис. 3.40 Взаимодействие
синаптических входов с гетерохимической организацией для стимулов
биологической (а) и сенсорных (б1 и б2) модальностей. Различные свойства входов
условно изображены виде размеров синапса и его активных зон..
|
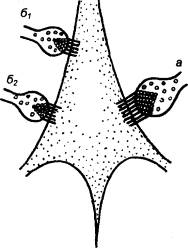
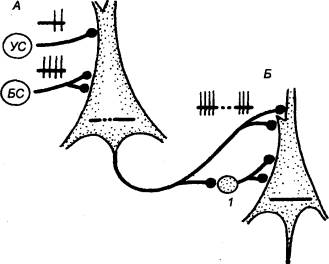
Клеточные механизмы условного
рефлекса.
Существуют две основные гипотезы научения: синоптическая, предполагающая,
что механизмом формирования условных рефлексов является изменение эффективности
синапсов, и мембранная, утверждающая, что основным механизмом условного
рефлекса является изменение свойств возбудимой постсинаптической мембраны.
Нервные механизмы научения также
обеспечиваются благодаря двум свойствам нервной системы: реактивности и
пластичности. Если реактивность — это способность отвечать на
раздражение, то пластичность — это способность изменять реактивность под
влиянием последовательных раздражении. Попытки использовать внутриклеточную
регистрацию постсинаптических электрических потенциалов привели к необходимости
проводить исследования либо на относительно простых нервных сетях, либо на
центральных нейронах высших млекопитающих с максимальным упрощением требуемых
для анализа межнейронных отношений. Такие упрощенные модели принято именовать клеточными
аналогами условного рефлекса. Эксперименты привели к выводу об определяющей
роли при выработке временной связи избирательных синаптических сдвигов.
Любой условный рефлекс начинается с
возникновения доминанты и суммационного рефлекса. Наиболее адекватным
отражением функционального состояния системы обучающихся нейронов является
уровень их мембранного потенциала, а следовательно, электротонические влияния
имеют наиболее существенное значение для образования временной связи. Отсюда
возникла поляризационно—электротоническая гипотеза формирования доминанты
и простых форм временных связей.
При использовании в качестве
ассоциируемых воздействий на клеточную мембрану подведение к ней биологически
активных веществ (глутамата и ацетилхолина), установлено различное проявление
пластичности для отдельных нейронов, вовлеченных в процесс ассоциации
раздражении. Показано, что в основе условной реакции втягивания жабр у морского
моллюска (аплизии) лежит механизм пресинаптического облегчения, который
формируется при конвергенции раздражителей, приводящих к спайковой активности
нейрона и активации этого же нейрона через модулирующий вход от безусловного
раздражителя. Аналогичные механизмы допускаются и у позвоночных животных.
Следовательно, и та, и другая гипотезы научения имеют своих сторонников.
Межнейронные отношения при
образовании временных связей. Исследования реакций нейронов бодрствующего мозга в
процессе научения показали, что наиболее существенные изменения характерны для
корковых полисенсорных нейронов, функциональные свойства которых
определяются диапазоном конвергирующих к ним афферентных влияний. Если до
первых сочетаний стимула с подкрепляющим рефлексом нейроны были преимущественно
моносенсорными, то по мере сочетаний они приобретают способность отвечать на
стимулы различных сенсорных модальностей и параметров, т. е. становятся
полисенсорными. Это соответствует периоду генерализации условного рефлекса со свойствами
доминанты. При достаточно упроченном условном рефлексе ответы нейронов снова
приобретают моносенсорный характер, но теперь это избирательная реакция только
на тот стимул, который приобрел сигнальное значение для поведенческого акта.
Возможно, что в процессе изменения
реактивности нейронов при образовании условного рефлекса определенное значение
имеют механизмы привыкания (пресинаптическая депрессия медиатора) и сенситизации,
или сенсибилизации (посттетаническая потенциация). Сенсибилизация в реагировании
нейронов коры мозга бодрствующей обезьяны возникает в том случае, если
раздражители прямо или косвенно связаны с мотивационной сферой животного.
Привыкание ответов тех же нейронов наблюдается в тех случаях, когда
используемые раздражители были индифферентными или хорошо знакомыми для
животного. Если ранее индифферентный агент приобретал свойства условного
сигнала, то ранее угашенные нейронные ответы возобновляются и стабилизируются
(рис. 3.42).
Те структуры, где изменения
происходят в первую очередь, рассматривают как наиболее существенные для
обучения. Критерием вовлеченности нейрона в условнорефлекторный процесс
является
|
|
|
Рис. 3.42 Динамика импульсной активности (а) и средней частоты, разрядов (в) коркового
нейрона лобной области обезьяны, а также электромиограмма мышц плеча (б) при
выработке защитного условного рефлекса А — при световом раздражении
(обозначено левой стрелкой); Б —
при кожном раздражении — укол (обозначено правой стрелкой); В, Г — при сочетании светового
раздражения с кожным. Приведенные реакции зарегистрированы при разном
количестве предъявленных условных раздражителей (А — 9 предъявлении, В — 1—9 сочетания, Г — 10—18 сочетания). |
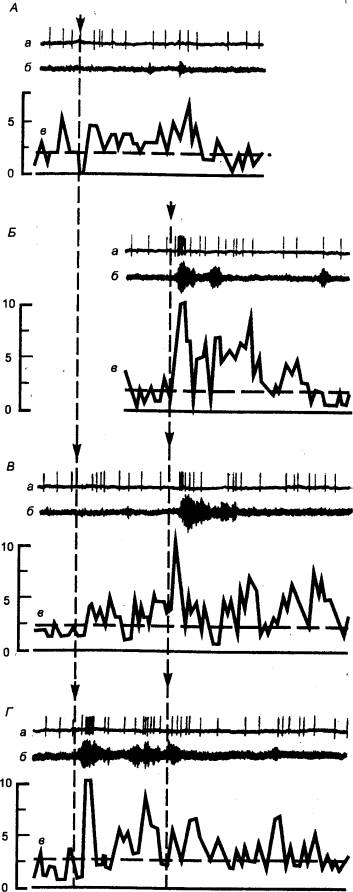
характер его предшествующей реакции,
из чего вытекает необходимость существования «критических синапсов», в которых
прежде всего происходят изменения, обусловливающие распространение импульсации
по новому пути.
В качестве наиболее значимых для научения
отделов мозга отмечены лобная и сенсорная кора, а также гиппокамп, где
обнаружены изменения в импульсной активности нейронов уже при первых сочетаниях
и упреждение их разряда поведенческой двигательной реакции. Реакции нейронов
сенсомоторной коры (снижение сопротивления мембраны и повышение возбудимости
клетки) предшествовали появлению первых условнорефлекторных движений и исчезали
при их отсутствии, хотя безусловнорефлекторная реакция сохранялась. Вероятно,
первичные пластические изменения происходят в нейронах коркового
представительства двигательной условной реакции. Эти перестройки могут
заключаться в изменении эффективности связей между нейронами сенсомоторной
коры, получающими возбуждение, и группами нейронов, контролирующих сокращение
определенных мышц'.
Выделяют элементарную
микросистему корковых нейронов при научении, приписывая ей следующие
основные свойства. 1. Деятельность микросистемы качественно отлична от
индивидуальной активности входящих в ее состав нейронов. 2. Системная
деятельность корковых нейронов на разных стадиях выработки условного рефлекса
основана на разных формах взаимоотношений между нейронами. 3. Микросистема
основывается на дифференцированном участии в ней различного класса нейронов,
по—разному меняющих свои сетевые свойства при научении. Таким образом,
определяющим механизмом временных связей оказываются не столько свойства самой
нервной клетки, сколько ее сетевые свойства как элемента системы.
Один из примеров возможных
межнейронных взаимоотношений в коре головного мозга при выработке условного
рефлекса приведен на рис. 3.43. Таламический афферент, вступающий в кору,
вызывает активацию тормозного интернейрона 3, непосредственно контактирующего с
эфферентной пирамидной клеткой 1. По мере выработки временной связи
данный нейрон начинает реагировать на условный сигнал той же тормозной
реакцией, как и на безусловный раздражитель. Ранее подпороговый вход условного
сигнала становится пороговым для запуска «безусловной» реакции. Но благодаря
участию в системе еще одного тормозного интернейрона 4, блокирующего проведение
импульсации от безусловного стимула к исследуемому нейрону, ответ нейрона 1
на безусловный стимул исчезает и возникает только по условному сигналу.
Нейрон обладает множеством входов в
составе сложнейших многонейронных конструкций, включающих много тормозных
интернейронов, однако наряду с механизмами локального облегчения синаптической
эффективности существует одновременно работающий механизм локального
подавления синаптических входов. В целом постсинаптическая мембрана
представляет собой сложную динамическую мозаику возбужденных и заторможенных
локусов, которые, будучи сочетанными во времени с активацией эндогенных
клеточных механизмов, фиксируются, обеспечивая вовлечение нейрона в состав общемозговой
нейродинамической системы.
Нейродинамическая констелляция
центров. А. А. Ухтомский
подчеркивал, что за каждой естественной доминантой кроется возбуждение целого
созвездия (констелляции) центров, своеобразная циклическая машина, функциональный
орган, образующийся по ходу самой реакции. Определяющим фактором при этом
является усваивание ритма и темпа возбуждения в центрах, входящих в состав
констелляции. М. Н. Ливанов (1972) разработал концепцию о пространственной
синхронизации
|
|
|
Рис. 3.43 Связи нервных клеток, объясняющие происхождение тормозных условных
нейронных ответов 1 — пирамидная клетка глубоких
слоев коры, 2 — звездчатая клетка, 3. 4 — тормозные интернейроны, 5 —
коллатерали аксонов, 6 — ветви апикальных дендритов в поверхностном слое
коры; ВС — путь проведения безусловного стимула (возбуждения) из таламуса, УС
— путь проведения условного стимула (возбуждения) от соседнего нейронного
модуля. |
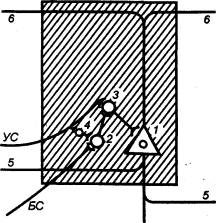
|
|
|
Рис. 3.44 Представление о связях в неокортексе на основании теории церебрального
научения Эклса Горизонтальные волокна аксонов ассоциативных (1) и комиссуральных (2) путей образуют синапсы с
апикальными дендри—тами звездчатых клеток в модуле Л и пирамидных клеток в
модулях Б и В. В модуле В показана шипиковая звездчатая
клетка (3) с аксоном (4),
образующим касательные синапсы (6)
со стволом апикального дендрита пирамидной клетки (5). |
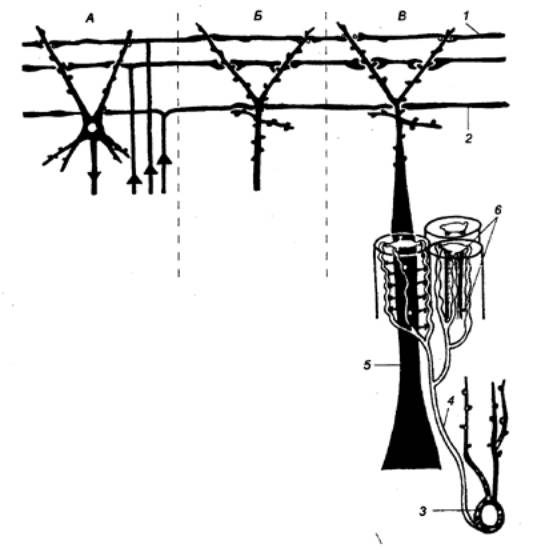
потенциалов отделов головного мозга как основе
формирования констелляции центров, ответственной за реализацию доминанты и ее условнорефлекторного
обеспечения. Эти представления получили развитие в гипотезе о распределенных
системах мозга, которая базируется на трех основных положениях. 1. Основные
структуры мозга построены по принципу повторения одинаковых многоклеточных
единиц — модулей. 2. Связи между основными образованиями мозга
многочисленны, избирательны и специфичны. 3. Каждая подгруппа модулей соединена
своей системой связей с такими же обособленными подгруппами в других отделах
мозга. Распределенные системы характеризуются избыточностью потенциальных
командных пунктов, а командная функция в разное время может локализоваться в
разных участках системы, особенно там, куда приходит самая срочная и нужная
информация.
Нейрофизиологические механизмы
сборки, формирования и поддержания столь обширных нейронных объединений в
единой рабочей системе приводит Дж. Эклс (1981). При этом исходными являются
три постулата. 1. Существование точной взаимной проекции модулей нескольким
обособленным группам модулей в том же полушарии и к зеркальному фокусу в
противоположном полушарии, что создает предпосылки для внутри— и межполушарной
реверберации. 2. Такие кольца связей существуют между корой и таламусом. Они
служат основой для таламо—корковой реверберации, в результате чего активируются
касательные синапсы на шипиках апикальных дендритов пирамидных клеток V слоя;
3. Апикальные дендриты пирамид образуют в I и II слоях касательные синапсы с
проходящими горизонтально аксонами.
Сочетание всех этих факторов
обеспечивает увеличение эффективности аксо—шипиковых синапсов, которые служат
основным звеном в формировании корковой пространственно—временной структуры
(констелляции) как матрицы памяти (рис. 3.44).
Действительно, изменение
геометрических параметров самого шипика меняет сопротивление для распространяющихся
по нему синаптических местных токов в направлении к стволу дендрита. Наряду с
жесткими структурными констелляциями существуют и гибкие звенья мозговых
систем, благодаря которым организм выполняет адаптивную деятельность в самых
разных условиях существования.
3.15.2. Механизмы условного
торможения
Каким образом неподкрепление условного сигнала приводит к ослаблению и прекращению условной реакции? Хотя И. П. Павлов и не давал однозначного ответа на этот вопрос, но высказывал предположение о том, что условное торможение возникает и первоначально локализуется в корковых клетках «представительства условного раздражителя». Другие ученые полагали, что условное торможение правильнее всего связывать с «корковым представительством безусловного рефлекса» или со структурами, осуществляющими временную связь между условным и безусловным раздражителями.
Трудно судить о приуроченности тех
или иных центральных процессов при условном торможении к работе строго
ограниченных структур мозга. Поэтому термин «внутреннее торможение» вряд ли
имеет строгое логическое обоснование в пределах системы тормозного условного
рефлекса.
При этом следует обратить внимание
на сходство закономерностей образования положительных и тормозных условных
рефлексов. Считается, что условный рефлекс возникает в результате синтеза двух
безусловных рефлексов, один из которых является ориентировочным на действие
условного раздражителя (рис. 3.45). Предполагается, что возбуждение
безусловного рефлекса производит индукционное торможение среднего звена дуги
ориентировочного рефлекса условного раздражителя. После прекращения действия
безусловного раздражителя данное торможение сменяется послетормозным
возбуждением. Последнее, в свою очередь, вызывает отрицательное индукционное
торможение условной связи. Это торможение охранительно—восстановительного типа
предотвращает следовое перевозбуждение условной связи и утомление
соответствующих корковых клеток. При отмене подкрепления нарушается
|
|
|
Рис. 3.45 Дуга
условного рефлекса с двусторонней связью А, Б — корковые
представительства мигательного и пищевого рефлексов; В, Г — подкорковые представительства мигательного и пищевого
рефлексов: 1,1' — афферентные
нейроны, 2, 2' — вставочные
нейроны, 3, 3' — эфферентные нейроны, 4,
5, 4', 5' — синаптические контакты коллатералей афферентных нейронов, 6 — синаптический контакт между
вставочным нейроном обратной условной связи и афферентным нейроном
сигнального раздражителя, 6' — синаптический
контакт между вставочным нейроном Прямой условной связи и афферентным нейроном
подкрепляющего раздражителя, I ,II — прямая и обратная условные связи. |
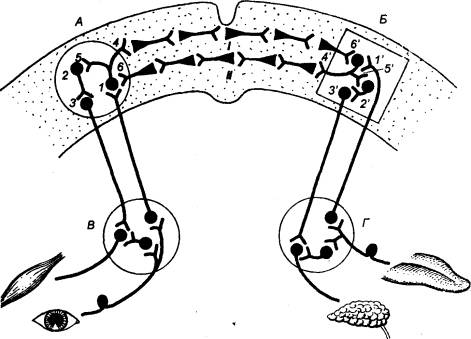
сопряженная индукция и элементы
условной связи испытывают длительное следовое возбуждение.
Согласно биологической гипотезе,
в основе механизма условного торможения лежит конкурентная борьба, столкновение
различных возбуждений. Более сильное возбуждение индукционно тормозит
выработанный пищевой рефлекс. Если при безусловном торможении такое возбуждение
возникает при ориентировочном или оборонительном рефлексе, то при условном
торможении — при биологически отрицательной реакции.
На рис. 3.46 приведена схема образования
угасательного торможения. Общий механизм тормозящего эффекта определяет единую
природу коркового торможения, в основе которого может лежать пессимальное, а
иногда и электротоническое торможение.
Считается, что при выработке
условного торможения формируется самостоятельный тормозный условный рефлекс,
независимый от положительного.
Приведенные гипотезы построены в
значительной мере на косвенных данных, а поэтому носят умозрительный характер,
нисколько не снижающий, впрочем, их оригинальной ценности.
Клеточный анализ условного
торможения показал, что в формировании тормозной реакции участвуют все звенья
положительного условного рефлекса, но они работают в другом режиме, меняются
параметры их деятельности. Отсутствие внешней реакции на отрицательный
(неподкрепляемый) стимул не связано с локальным или системным торможением
структур головного мозга. В ответ на тормозное раздражение возникает такая же
многообразная активность нейронов, как на положительное раздражение.
Некоторые исследователи видят
проявления условного торможения в снижении или прекращении импульсной
активности клетки при продолжительном действии положительного условного сигнала
(торможение запаздывания). Обнаружено, что при угасательном торможении
амплитуда ВПСП постепенно снижается без заметного перехода в гиперполяризацию
клеточной мембраны. Появилась возможность объяснить ослабление возбуждающих
реакций уменьшением выброса медиатора без активного торможения самого нейрона.
Подобные наблюдения, а также факты постепенного угнетения клеточных реакций при
угашении могут быть связаны с понижением возбудимости корковых
|
|
|
Рис. 3.46 Образование угасательного торможения А — условный пищевой рефлекс; Б — стадия частичного
торможения пищевой реакции вследствие появления биологически отрицательной
реакции; В — стадия полного торможения пищевой реакции: / — анализатор, 2 — корковое представительство
пищевой реакции, 3 — корковое представительство биологически отрицательной
реакции, 4— путь для
побочного торможения |
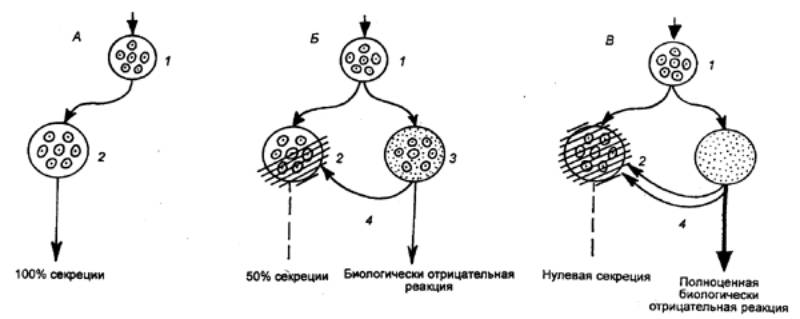
нейронов. Вероятно, при развитии
угасательного торможения имеет место размыкание временной связи.
Положительные и отрицательные
условные рефлексы опосредуются разными синаптическими входами активирующего и
тормозного действия. В зависимости от того, какие интернейроны включены в
полисинаптические пути условной связи, дифференцировочное торможение будет
проявляться или как угнетение активирующих условных реакций клетки, или как
растормаживание клеточных реакций с тормозной условной реакцией.
Самостоятельная группа клеток на неподкрепляемое раздражение усиливала
импульсацию, а на подкрепляемый положительный сигнал не реагировала вовсе.
При выполнении инструментальных
условных рефлексов на комплексный сигнал вырабатывали условное торможение
реакций при изолированном предъявлении компонентов (рис. 3.47). Установлено,
что паттерн ответа этих нейронов на положительный сигнал (свет + звук)
существенно отличался от такового при дифференцировочных сигналах (свет, звук).
Эти паттерны ответов обусловлены двигательной программой, которая реализуется
по данному сигналу. Если угасательное торможение сопровождается распадом
паттерна нейронных ответов и/или их исчезновением, то дифференцировочное
торможение, тренируемое вместе с положительным рефлексом, стабильно
поддерживает новый паттерн нейронного ответа, отражающий центральную программу
поведенческой реакции.
При разных видах условного
торможения существует три типа перестроек импульсации: 1) инверсия
паттерна активности нейрона, имеющая место при положительном сигнале, в
противоположный по знаку при предъявлении отрицательного сигнала; 2)
рекомбинация паттерна ответа при предъявлении неподкрепляемого раздражителя в
сравнении с эффектом положительного сигнала; 3) исчезновение реакции нейронов
при предъявлении дифференцировочного раздражителя. Эти перестройки именуются
как центральный нейронный эквивалент дифференцировочного торможения.
Условное торможение является таким
же активным системным процессом, как и условное возбуждение. По своей
организации, за исключением конечного эффекторного звена, оба процесса подобны.
Вероятно, мозаика постсинаптических
процессов, происходящих на корковом уровне, отражает перестройки функциональной
активности соответствующих центральных программ, эффекторное выражение которых
осуществляется на соответствующих
|
|
|
Рис. 3.47 Перистимульная гистограмма активности нейрона сенсомоторной
коры кошки при положительных и отрицательных условных рефлексах А — положительные условные
рефлексы на комплексный сигнал из одновременных светового и звукового
компонентов; Б, В — отрицательные условные рефлексы
на изолированные предъявления светового и звукового компонетов. По оси
ординат — число импульсов, каждый столбик соответствует периоду в 50 мс, по
оси абсцисс — время в секундах, каждая гистограмма построена на основе 10
реализации; под гистограммой: отклонение вверх — условный сигнал, вниз —
подача подкрепления, горизонтальные отрезки — проявление двигательной реакции
передней конечности. |
|
|
|
Рис. 3.48
Временная организация памяти. (Объяснение см. в тексте). |
Основными свойствами нервной системы
являются длительное хранение информации о событиях внешнего мира и реакциях
организма на эти события, а также использование этой информации для построения
текущего поведения. Эти свойства легли в основу формирования нервной,
или нейрологической, памяти. В каждый момент времени животные и человек
переходят из своего прошлого к новому ранее не испытанному состоянию, и их
поведение в настоящем в значительной мере определяется совокупностью
накопленного опыта в прошлом. Память как результат научения заключается в таких
изменениях в нервной системе, которые сохраняются в течение некоторого времени
и существенно влияют на характер протекания будущих рефлекторных реакций.
Комплекс таких структурно—функциональных изменений получил название процесса
образования энграммы (от греч. en— находящийся внутри и gramma— запись) — материальной
основы памяти.
Память характеризуется системной организацией, т. е.
изучение элементарных механизмов памяти должно ориентироваться на целостное
понимание этой функции мозга. Известно, что в памяти события размещаются по оси
времени с отражением их реальной длительности. Формирование и воспроизведение
таких энграмм возможно лишь при допущении временного «свертывания» событий на
основе существования собственного времени мозга. В результате такой компрессии
времени создается внутренний хронотоп, т. е. внутренний
пространственно—временной образ внешнего единого мира. Энграмма из модели
прошлого перемещается в будущее и становится фактором, определяющим цель
предстоящего поведенческого акта.
Временная организация памяти. После электрического ответа
рецепторной клетки на внешнее воздействие возникают следовые процессы,
продолжающиеся при отсутствии реального раздражителя. Они и составляют основу
сенсорной памяти. Длительность хранения следов в сенсорной памяти не
превышает 500 мс, стирание следа осуществляется за 150 мс. Сенсорная память
человека не зависит от его воли и не может быть подвергнута сознательному
контролю. Следующий период, связанный с хранением информации, именуется кратковременной
памятью. И наконец, наиболее значимая информация хранится в долговременной
памяти. Консолидация энграммы, т. е. переход ее из кратковременной в
долговременную, соответствует особому виду промежуточной памяти.
Предполагают, что кратковременная (нестабильная) память
образуется за счет конформационных перестроек макромолекул и реализуется на
синаптическом и синаптосомальном уровнях регуляции (рис. 3.48). Перемещение
ионов и/или кратковременные метаболические сдвиги во время синаптической
активности могут привести к изменению эффективности синаптической передачи,
длящейся миллисекунды и секунды.
Большинство исследователей полагают,
что в основе кратковременной памяти лежат электрофизиологические механизмы,
связанные с реверберацией импульсации по замкнутой системе нейронов.
Синаптосомальный
уровень включает конформационные изменения структурных и ферментных белков, а
также перемещение нейромедиаторов — длящаяся минуты и часы промежуточная память.
Долговременная память формируется на основе синтеза макромолекул (нуклеиновых
кислот и белков) и связана с активацией генетического аппарата.
В отличие от
предшествующих процессуальных видов долговременная память представляет собой
новую внутримозговую функциональную структуру, базирующуюся на изменениях в
мембранах нейронов и межнейронных связях.
3.16.1. Кратковременная память
В качестве наиболее удачной модели
изучения кратковременной памяти используют отсроченные условнорефлекторные
реакции, состоящие в том, что между условным сигналом и соответствующим
двигательным ответом вводится пауза (отсрочка), после окончания которой
животное получает доступ к рычагам и кормушкам. Правильность выполнения
двигательного акта свидетельствует об удержании в кратковременной памяти
информации об условном сигнале. Особенности импульсной активности нейронов в
период отсрочки могут быть использованы для понимания механизмов краткосрочной
памяти.
При этом особый интерес представляют лобные доли больших
полушарий, часть которых у обезьян именуется «фокусом пространственной памяти».
Исследования на обезьянах показали, что время отсрочки заполняется краткими
вспышками импульсивной активности нейронов. Нейроны, активирующиеся в начальный
период отсрочки, детектируют информацию о свойствах условного сигнала, далее
следует передача возбуждения по принципу эстафеты другим нейронным популяциям,
каждая из которых отдает аксоны к нейронам — сумматорам, и от последних
происходит передача задержанной таким образом информации на программирующие и
пусковые структуры мозга (рис. 3.49).
|
|
|
Рис.
3.49 Основные типы.
перестроек импульсной активности нейронов лобной коры обезьяны (А—Д) в период
краткосрочной памяти (отсрочка) По оси ординат — активность, имп/200
мс. Стрелками указаны начало и окончание отсрочки, цифрами на кривых —
процентное соотношение нейронов каждого типа. |
Для лобных отделов коры характерны замкнутые нейронные круги
типа «ловушек», в которых импульсный поток может реверберировать до тех пор,
пока не будет переключен на эфферентные корковые модули из крупных пирамидных
клеток (рис. 3.49). Теменная кора, также участвующая в кратковременной памяти,
снабжена механизмом таламо—корковой реверберации, которая в отличие от лобной
коры продолжается на протяжении всего периода отсрочки.
Считается, что фиксация следового
процесса связана со стойкими изменениями синаптической проводимости,
возникающими при повторном поступлении к синапсу циркулирующего импульсного
потока определенной конфигурации. В составе замкнутых нейронных цепей в коре
мозга участвуют тормозные интернейроны, которые способствуют более длительному
хранению следов и поддержанию циркулирующей активности.
Среди многих гипотез памяти
рассмотрим представление, согласно которому ионные сдвиги в результате
упражнения (научения) синапсов запускают длительные восстановительные процессы.
Следствием их является гиперполяризация мембраны, которая сопровождается
поступлением аминокислот и синтезом некоторых белков.
Вновь синтезированный белок
используется синапсами, стабильно меняющими свою эффективность. При этом
указываются следующие временные границы видов памяти: кратковременная — угасает
через 10 мин после научения, промежуточная— возникает в течение 15 мин после
научения и угасает через 30 мин; долговременная — возникает через 45 мин после
научения и сохраняется неопределенно долгое время.
Классификация видов памяти. И. С. Бериташвили (1975) различал
образную, условнорефлекторную, эмоциональную и словесно—логическую память. Под образной
памятью человека и животных понимают сохранение в памяти и репродукцию
однажды воспринятого жизненно важного объекта; под эмоциональной памятью —
воспроизведение пережитого ранее эмоционального состояния при повторном
воздействии раздражителей, обусловивших первичное переживание этого состояния.
Эмоциональная память обладает следующими особенностями: 1)
она надмодальна, т. е. ее формирование и воспроизведение может происходить при
любых сенсорных воздействиях; 2) она реализуется очень быстро и часто с первого
раза; 3) она характеризуется непроизвольностью запоминания и воспроизведения
информации, т. е. обеспечивает пополнение подсознательной сферы человеческой психики.
Условнорефлекторная память проявляется в виде воспроизведения условных
двигательных и секреторных реакций или заученных привычных движений спустя
длительное время после их образования.
Наконец, словесно—логическая память — это память на
словесные сигналы, обозначающие внешние объекты, события, внутренние
переживания и свои собственные действия.
В любом случае происходит фиксация образов, предметов и
явлений, включающая в свой состав по крайней мере три этапа: формирование
энграммы, сортировку и выделение новой информации, долговременное хранение
значимой для организации информации.
Основная гипотеза базируется на признании того, что фиксация
следа памяти связана со стойкими изменениями синаптической проводимости
в пределах определенного нейронного ансамбля.
В основе долговременной памяти лежат сложные
структурно—химические преобразования на системном и клеточном уровнях головного
мозга.
Под влиянием научения увеличивается количество
холинорецепторов, что может быть следствием либо синтеза новых рецепторных
молекул, либо демаскировки и активации уже существующих рецепторных белков.
Установлено повышение чувствительности кортикальных нейронов к ацетилхолину в
процессе образования условного рефлекса. Активация рецепторов постсинаптической
мембраны облегчает научение, ускоряет фиксацию, способствует извлечению следа
из памяти. И наоборот, антагонисты ацетилхолина нарушают научение и
воспроизведение, вызывая амнезию (потерю памяти).
Катехоламинергическая и
серотонинергическая системы принимают самое непосредственное участие в
механизмах долговременной памяти. Установлено, что научение животных в моделях
с электрокожным подкреплением условных рефлексов сопровождается активацией
адренергических систем мозга, а научение с пищевым подкреплением — снижением
метаболизма и уровня норадреналина в головном мозгу. '
Предполагают, что норадреналин
пролонгирует активность нейронов, вызванную предъявлением условного стимула, и
этим облегчает формирование условного рефлекса. Серотонинергической
системе принадлежит важная роль в процессах консолидации следов памяти.
Серотонин ускоряет научение и удлиняет сохранение навыков, выработанных на
эмоционально положительном подкреплении, нарушая выполнение и сохранение
защитно—оборонительных реакций. Согласно существующей концепции, моноамины
участвуют в процессах научения и памяти опосредованно, через нейрохимическое
обеспечение положительных и отрицательных эмоций. Серотонин причастен к
формированию эмоционально положительных, а норадреналин — эмоционально
отрицательных состояний.
Существует гипотеза, согласно
которой холинергические механизмы мозга обеспечивают информационную
составляющую процессов научения. Роль моноаминергических систем мозга
сводится к облегчению подкрепляющих и эмоционально—мотивационных составляющих
процесса научения и памяти, причем холинергическая система находится под
модулирующим влиянием моноаминергической системы. Стимуляция моноаминергических
механизмов перестраивает хемореактивные свойства, которые при определенных
условиях могут закрепляться, обеспечивая создание и сохранение многонейронной
констелляции — энграммы. Если адренергические механизмы мозга в большей мере
причастны к формированию временных связей, то серотонинергические — к их
фиксации.
Процессы долговременной памяти
зависят также от уровня метаболизма ГАМК, глутаминовой кислоты, вторичных
посредников (циклических нуклеотидов и ионов кальция).
Участие информационных макромолекул
(нуклеиновых кислот и белков) в ключевых процессах научения и памяти не
вызывает сомнений. Согласно одной группе гипотез, научение и память связаны с
кодированием приобретенных форм поведения в информационных макромолекулах.
Согласно другой группе гипотез, которая исходит из взаимосвязи генома и синтеза
специфических белков нервной клетки, на основе функционального объединения
нейронов возникает структурное их объединение, представляющее собой энграмму
памяти.
Первая группа гипотез базируется на
аргументах о качественном изменении РНК и белков при научении и возможности
переноса памяти от обученного мозга к необученному с помощью РНК или
полипептидов. Установлено, что для процессов консолидации и формирования
долговременной памяти необходим полноценный белковый синтез. В формировании
устойчивости созданной энграммы должна существовать система обновления
специфических рецепторных белков либо должны возникать стабильные модификации
ДНК, в
|
|
|
Рис.
3.50 Основные типы
нейронных объединений в лобной коре обезьян А — нейронная «ловушка»; Б — вертикальный модуль. |
|
|
|
Рис. 3.51 Аксошипиковый транзиторный синапс 1 — аксон, 2 — синаптические везикулы, 3 — шипиковый аппарат, 4 — нейроглия, 5 — молекула актина,
б — рибосомы, 7 — митохондрия, 8 —
дендрит, 9 — дендритные
трубочки; I — влияние
пресинаптического возбуждения на постсинаптическую мембрану, вызывающее в головке
шипика возрастание концентрации Са2+; II — влияние Са2+ на
молекулы актина, приводящее к сокращению ножки шипика и облегченному
проведению местного потенциала к стволу дендрита. |
результате которых в нейронах образуется
и поддерживается пожизненно синтез любого нейроспецифического белка или,
наоборот, необратимо выключается синтез маскирующего белка.
Полагают, что повторная импульсация в нейроне сопровождается
увеличением концентрации кальция в постсинаптической мембране. Это активирует
фермент — кальций—зависимую протеиназу, которая расщепляет один из
белков мембраны. Его расщепление высвобождает замаскированные ранее неактивные
белковые глутамат—рецепторы. С возрастанием числа активных глутаматрецепторов
возникает состояние повышенной проводимости аксошипикового синапса (в течение
3—6 сут).
Аксошипиковые контакты — наиболее пластичное соединение между нейронами,
которое может быть ответственно за эффективность синаптической проводимости.
Последнее достигается изменением диаметра самого шипика, что меняет, в свою
очередь, сопротивление мембраны. Это обеспечивается наличием в ножке шипика
контрактильного аппарата в форме молекул актомиозина. Их активация возникает
при высвобождении ионов кальция из депо, коим является расположенный в головке
шипиковый аппарат (рис. 3.50, 3.51). Высвобождение ионов кальция происходит при
действии медиаторов на постсинаптическую мембрану. Сокращение молекул
актомиозина приводит к укорочению и утолщению ножки шипика, вследствие чего меняется
сопротивление и проведение электрического потенциала к дендритному стволу.
Для понимания механизмов «переноса памяти» целесообразно
использовать гипотезу И. П. Ашмарина об участии иммунологических механизмов
в долговременной памяти (рис. 3.52). Если представить, что после
прохождения импульсов через синапс усиливается синтез специфических
белков—антигенов, то их избыток должен
|
|
|
Рис.
3.52 Представление
об иммунохимическом механизме формирования памяти (по Ашмарину) КБ — катионные белки, открывающие путь комплексам
синаптического антигена с РНК через оболочку клетки; Л — клетка глии (аналог
лимфоцитов); М — клетка глии (аналог макрофагов); 1, 2, ? — участки синаптического антигена (зона синтеза
заштрихована); 4 — комплекс
фрагмента синаптического антигена с РНК; 5 — антитела для синапсов. Тонкими
линиями изображены мРНК, толстыми — ДНК хромосом,. |
выходить в околосинаптическое
пространство. Эти белки взаимодействуют с рядом расположенными клонами клеток астроцитарной
глии, индуцируют их размножение и образование антител. Последние специфически
взаимодействуют с постсинаптическими мембранами тех же нейронов и облегчают
проводимость в соответствующих синапсах. Данный клон астроцитов сохраняется в
течение жизни. Следовательно, действующим началом «переноса памяти» может быть избыточный
антиген пептидной природы, который способен автоматически найти в мозгу
реципиента либо соответствующую клетку глии либо синапс.
'
Специальные исследования на животных с генетическим
дефицитом вазопрессина и нарушением памяти обнаружили ее улучшение после
инъекции дополнительных количеств этого гормона. Противоположное действие —
нарушение сохранения выработанных навыков — оказывает другой гормон гипофиза — окситоцин.
Эндорфины и энкефалины (эндогенные опиаты) оказывают выраженное
влияние на научение и память — замедляют угашение
условных
рефлексов, улучшают их сохранение, хотя и ухудшают их формирование.
Нейропептиды обнаружены в аксонных
окончаниях нейронов вместе с классическими медиаторами. Нейропептиды могут
усилить или ослабить действие медиатора. Нейропептиды—спутники вместе с
медиаторами создают и поддерживают на постсинаптической мембране специфические
рецепторные мозаичные наборы, способствующие быстрому проведению определенного
вида возбуждения. Пептид—спутник повышает сродство рецептора к основному
медиатору, он более стабилен, чем основной медиатор, что обеспечивает
пролонгированное облегчение проведения через синапс.
Вероятно, все формы нейрологической
памяти обеспечиваются системой межнейронных взаимодействий, участие же
информационных молекул проявляется лишь во включении или выключении различных
участков предсуществующего генома, а не в синтезе новых нуклеотидных или
аминокислотных последовательностей.
3.17. ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА И ПОВЕДЕНИЕ
Принцип доминанты в
качестве основы складывающихся координационных отношений выдвинул А. А.
Ухтомский. Он активно опирался на идеи Ч. Шеррингтона о конвергенции афферентных
влияний к аппаратам, образующим общие пути в ЦНС. Но лишь идеи Н. Е.
Введенского о динамизме взаимоотношений в нервной системе, обусловленные
характером воздействия, функциональным состоянием нервных центров и
характеристиками рабочего органа, позволили сформулировать учение о доминанте —
рабочем принципе нервной деятельности и векторе поведения.
3.17.1. Доминанта и условный рефлекс
Доминанта как скрытая
предуготовленность организма к выполнению конкретного поведения определяет
вероятность возникновения той или иной рефлекторной реакции (условной или
безусловной) в ответ на текущее раздражение.
Еще И. М. Сеченов обращал внимание
на то, что иррадиация возбуждения происходит в сторону очагов повышенной
возбудимости. Определяющую роль при формировании такого очага, по Ухтомскому,
играет состояние развивающегося в нем стационарного возбуждения. Если
этот уровень возбуждения мал, то приходящие сюда нервные влияния могут поднять
его до состояния, характерного для доминанты, т. е. создать в нем повышенную
возбудимость. Если же уровень возбуждения в центре и без того высок, то при
приходе новой волны возбуждения возникает эффект торможения.
Важными особенностями доминантного
очага являются повышенная возбудимость, т. е. отзывчивость на приходящие
волны возбуждения, и способность центра суммировать эти возбуждения. В
доминантном очаге устанавливается определенный уровень стационарного
возбуждения, способствующий суммированию ранее подпороговых возбуждений и
переводу на оптимальный для данных условий ритм работы, когда этот очаг
становится наиболее отзывчивым.
Следовательно, доминанта, прежде чем
стать таковой, должна пройти стадию суммационного рефлекса. В то же время
условный рефлекс, прежде чем стать таковым, проходит стадию доминанты (II
стадия генерализованной, диффузной отзывчивости).
Важную роль в процессе формирования
доминанты играет процесс сопряженного торможения. Состояние возбуждения
в центре, подкрепленное возбуждениями из самых отдаленных источников, будучи
достаточно стойким, т. е. инертным, в свою очередь снижает способность
некоторых других центров реагировать на импульсы, имеющие к ним прямое
отношение. Сопряженное торможение должно быть своевременным, т. е. иметь
координирующее значение для работы других органов и организма в целом. Это наиболее
энергетически емкий процесс, наиболее тонкий, позднее вырабатывающийся и вместе
с тем наиболее ранимый.
Процесс торможения не только
оформляет и поддерживает доминанту, но может и привести к прекращению ее
функционирования, т. е. сама доминанта может тормозиться. А. А. Ухтомский
выделял четыре случая торможения доминанты: 1) если доминанта
представляет собой цепной рефлекс, то разрешающий акт и будет концом доминанты
(например, половое поведение); 2) возникновение в центрах новой доминанты,
несовместимой с первой, ведет к ее торможению (защитное поведение тормозит
пищевое); 3) осуществление прямого торможения доминанты возможно волевым путем,
т. е. из коры мозга (подавление естественных физиологических потребностей в
неадекватных условиях); 4) стойкое подкрепление доминанты посторонними
импульсами само по себе может подготовить ее торможение (доминанта в самой себе
несет свой конец, обладая способностью растормаживаться).
В существовании доминанты большую
роль играет временной фактор. Именно на длительности функциональных
сдвигов в центрах и базируются такие свойства доминанты, как стойкость,
инертность, способность к суммации. Доминанта опирается на наличие
определенного набора оптимальных раздражителей, но в основном на следовые
процессы, которые и отличают ее от условных и безусловных рефлексов (частных
коротких рефлексов, по Ухтомскому).
|
|
|
Рис. 3.53 Соотношение первичных и вторичных
очагов возбуждения при формировании доминирующей констелляции |
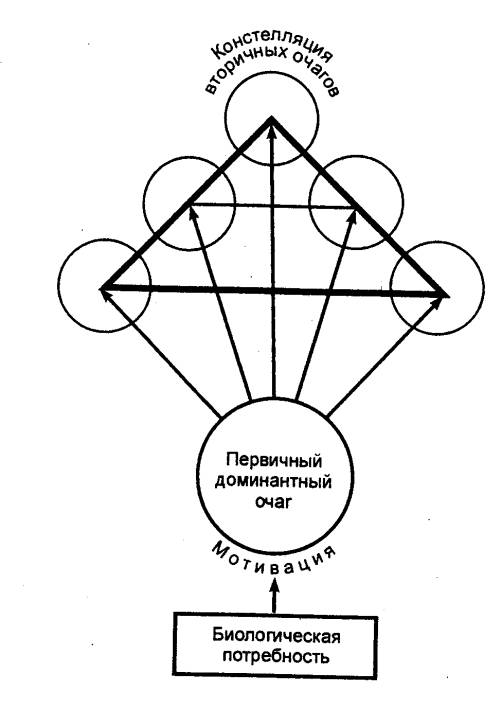
Сложные рефлекторные акты осуществляются
не одним анатомическим центром, а несколькими, которые как бы образуют
своеобразную функциональную констелляцию центров в работе целостного мозга.
Каждый участник этой констелляции, будучи связан с соседними и стимулируя их, в
свою очередь получает от них стимуляцию. Такое состояние было названо симптомокомплексом
доминанты.
Первичный очаг возбуждения при
формировании естественных биологических доминант (голода, жажды) возникает в
структурах гипоталамуса, содержащих большое число специфических хеморецепторных
нейронов. Этот очаг выступает в качестве важнейшего системообразующего фактора
при формировании доминирующей констелляции, в которую входит множество
вторичных очагов в тех отделах мозга, где существуют предпосылки для
продолжительного удержания следовых процессов (гиппокамп, кора больших
полушарий) (рис. 3.53).
Для понимания механизмов превращения доминанты из
симптомокомплекса А.А. Ухтомский использовал понятие функционального центра,
или органа. Этим подчеркивалось следующее: дело не столько в том, что в
нервной системе складываются определенные констелляции связанных между собой
центров, морфологически далеко разбросанных по мозговой массе, но и в том что
они функционально объединены единством действий, своей векторной направленностью
на определенный результат.
В отличие от технического
механизма, такой функциональный орган образуется по ходу самой реакции и имеет
множество степеней свободы, дающих ему возможность осуществлять и множество
переменных операций. Единство их действия достигается способностью данной
совокупности мозговых структур влиять друг на друга с точки зрения усвоения
ритма, т. е. синхронизации их активности. Каждый из потенциальных компонентов
констелляции изначально обладает своим определенным темпом и ритмом возбуждений.
При формировании же единой констелляции как созвездия созвучно работающих
ганглиозных участков, совозбуждающих друг друга, определяющую роль играет
увязка во времени, скоростях и ритмах действия, т. е. в сроках выполнения
отдельных моментов реакции. Доминирование данной констелляции будет зависеть от
синхронизации активности входящих в нее компонентов и от сопряженного
торможения других рефлексов.
В своем формировании рабочая констелляция проходит несколько
стадий. 1. Вначале в констелляцию посредством генерализованного возбуждения
наряду с необходимыми для данного акта нервными центрами вовлекаются и другие
мозговые структуры. Доминанта вызывается в центрах непосредственным
раздражителем, часто гуморальной природы, или рефлекторными влияниями. На этой
стадии она и подкрепляется самыми разнообразными внешними раздражениями и
уподобляется стадии генерализации условного рефлекса. 2. Постепенно в процессе
повторного осуществления данного рефлекторного акта такая диффузная
отзывчивость сменяется избирательным реагированием только на те раздражения,
которые создали данную доминанту. А. А. Ухтомский назвал это стадией
образования условного рефлекса, когда подбирается адекватный раздражитель
для данной доминанты. И именно эти раздражители способны воспроизвести прежнюю
доминанту со свойственными для нее вегетативным и соматическим проявлениями.
Продукт пережитой доминанты, ее своеобразная памятная энграмма, в которой в
единое целое сплетены поведенческие, мотивационно—эмоциональные ее признаки и
комплекс вызывавших ее раздражении, были названы А. А. Ухтомским интегральным
образом. Эта стадия доминанты строится на законе «обратных отношений», по
которому репродуцируется доминанта при наличии адекватных сигналов.
Жесткие рабочие констелляции характеризуются непременными
генетически запрограммированными результатами выполнения рефлекторного акта.
Кора больших полушарий формирует новые констелляции в индивидуальной жизни
организма. Такие кортикальные доминанты обеспечивают организму при всем
разнообразии конкретной среды определенный и устойчивый вектор поведения.
Воспроизведение доминанты по ее кортикальным следам может быть весьма
экономичным (эскизным), и, комплекс органов, участвующих в переживании
восстановленной доминанты, может быть очень сокращенным. Каждый раз воспроизведенный
интегральный образ обогащается все новыми элементами, что служит основой для
расширения и перестройки индивидуального жизненного опыта.
Таким образом, кора больших полушарий формирует адекватные
стимулы для доминанты, запускает соответствующие доминанты по поводу этих
стимулов и обеспечивает взаимодействие и переключение доминант. Такие свойства
в наибольшей мере присущи ассоциативным корковым полям. Следовательно, синтез
механизмов доминанты с механизмами образования условного рефлекса необходим и
достаточен для организации целенаправленного поведения. Доминанта обеспечивает
его активный творческий характер, а упроченный специализированный условный
рефлекс — его адаптивный характер, т. е. точное соответствие поведения
объективной реальности.
3.17.2. Высшие интегративные системы мозга
Наряду со
специфическими и неспецифическими системами в самостоятельную категорию
выделяют ассоциативные таламокортикальные системы. Применительно к
высшим млекопитающим это своеобразные структуры, не принадлежащие к какой—либо
одной сенсорной системе, но получающие информацию от нескольких сенсорных
систем. Ассоциативные ядра таламуса относятся к «внутренним ядрам», афферентные
входы к которым идут не от сенсорных специфических путей, а от их
переключательных образований. В свою очередь, эти ядра проецируются на ограниченные корковые
территории, именуемые ассоциативными полями.
Согласно анатомическим данным,
различают две ассоциативные системы: таламопариетальную и таламофронтальную.
Среди таламических ядер, проецирующихся на теменную кору, по
многообразию связей и особенностям нейронной активности особое место занимает
комплекс заднее (каудальное) латеральное ядро — подушка таламуса. Сама
теменная кора является местом широкой гетеросенсорной конвергенции по путям от
специфических, ассоциативных и неспецифических ядер таламуса, а также по
волокнам от сенсорных корковых зон и симметричной коры противоположного
полушария.
Таламопариетальная
ассоциативная система
представляет собой: 1) центральный аппарат первичного одновременного анализа и
синтеза обстановочной афферентации и запуска механизмов ориентационных движений
глаз и туловища; 2) один из центральных аппаратов «схемы тела» и сенсорного
контроля текущей двигательной активности; 3) важнейший аппарат предпусковой интеграции,
участвующий в формировании целостных полимодальных образов.
Таламофронтальная
ассоциативная система
представлена медиодорсальным ядром таламуса, проецирующимся на лобную
долю больших полушарий. Последнюю, имеющую множество нисходящих связей, рассматривают
в качестве коркового модулятора лимбической системы. Основная функция
таламо—фронтальной системы состоит в программировании целенаправленных
поведенческих актов на основе доминирующей мотивации и прошлого жизненного
опыта.
В качестве основных механизмов
работы ассоциативных систем определяют следующие.
1.
Механизм
мультисенсорной конвергенции — к
ассоциативным полям коры конвергируют афферентные посылки, несущие информацию о
биологической значимости того или иного сигнала. Разделенные афферентные
влияния вступают в интеграцию на кортикальном уровне для формирования программы
целенаправленного поведенческого акта.
2.
Механизм пластических
перестроек при гетеромодальных сенсорных воздействиях, которые проявляются либо в избирательном привыкании,
либо в сенситизации, либо в формировании ответов экстраполяционного типа.
Доминирующая мотивация определяет спектр конвергирующих модальностей и характер
их корковой интеграции.
3. Механизм краткосрочного хранения
следов интеграции, заключающийся в
длительной внутрикорковой или таламокорковой реверберации импульсных потоков.
3.17.3. Эволюция ассоциативных
систем
Развитие
современных млекопитающих шло параллельными рядами, и хотя общий план
конструкции мозга сохранился, таламокортикальные системы претерпели наиболее
существенные морфофункциональные перестройки. Рассматривая степень развития
ассоциативных систем как показатель филогенетического статуса вида, можно
выделить три основных уровня их эволюции (рис. 3.54).
Мозг
насекомоядных рассматривают как предшественник с его дальнейшим усложнением в
параллельных рядах эволюции грызунов, хищных и приматов. У насекомоядных
выделена примитивная ассоциативная таламокортикальная система, которая хотя и
участвует в процессах сенсомоторного синтеза т е непосредственной сенсорной
активации выходных элементов коры, но не способна к формированию более сложных
актов сенсорной интеграции модально специфических влияний, предшествующих
запуску выходных корковых нейронов.
|
|
|
Рис. 3.54 Эволюционное созревание
интегративных аппаратов мозга у насекомоядных (А), хищных (Б) и приматов (В)
Выделены лишь две специфические сенсорные системы — зрительная (1) и соматическая (2); 3 — таламус, 4 — кора, 5,6 — таламофронтальная и таламопариетальная ассоциативные
системы больших полушарий; светлыми линиями обозначены проекционные пути
зрительной и соматической сенсорных систем; заштрихованы — ассоциативные
системы; черными сплошными стрелками — кортико—кортикальные связи,
пунктирными — эфферентные корковые проекции (преимущественно пирамидный
тракт). |
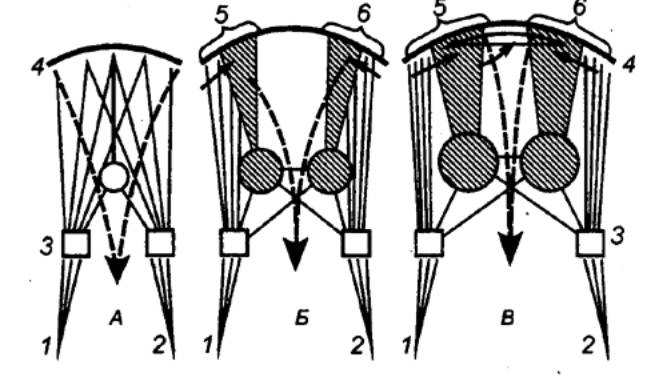
У грызунов, мозг которых близок к
насекомоядным, слабая выраженность морфологической дифференциации и
функциональной специализации полисенсорных структур определяет несовершенство
интегративной деятельности мозга.
У хищных впервые появляются в
коре больших полушарий развитые и относительно автономные лобные и теменные
ассоциативные поля и соответствующие структуры таламуса. Характерны структурные
и функциональные различия ассоциативных систем от других мозговых структур и
между собой. Таламопариетальная система — следствие усложнения конструкции и
связей зрительной сенсорной системы — участвует в сложных актах
пространственной ориентировки и обеспечивает текущий сенсорный фон для
выполнения целенаправленных поведенческих актов. Таламофронтальная система
включается в своей значительной части в корковый отдел скелетно—мышечной
сенсорной системы с одновременной проекцией на нее лимбических структур. Она
участвует в организации программ целостных двигательных актов. В пределах
отряда хищных лобные поля неокортекса усложняются, увеличиваются их размеры и
роль в поведении, требующем мобилизации механизмов кратко— и долговременной
памяти.
Перекрытие с зонами выхода
эфферентных корковых путей, достаточная роль докоркового уровня интеграции,
преобладание в каждой ассоциативной системе какого—либо одного из сенсорных
входов не способствуют достижению полной гетеросенсорной интеграции.
У приматов ассоциативные
структуры таламуса с их обширной и дифференцированной проекцией в лобные и
теменные области коры образуют целостную интегративную систему больших
полушарий. Это достигается прежде всего с помощью развитых кортико—кортикальных
ассоциативных связей. Благодаря компактной системе миелинизированных пучков
волокон возрастает роль кортикального уровня взаимодействия специфических
сенсорных зон с ассоциативными полями. Последние характеризуются тонкой
дифференциацией с формированием из нейронных элементов целостных
структурно—функциональных ансамблей (модулей).
Функциональная значимость
ассоциативных систем приматов расширяется и уточняется по сравнению с хищными.
Утрачивается преобладание какого—либо одного сенсорного входа, а следовательно,
расширяются возможности их интеграции. Возникает топографическая разнесенность
ассоциативных полей от собственно эфферентных корковых формаций, что снижает
удельное значение сенсомоторной интеграции и расширяет роль коры в
осуществлении межсенсорного афферентного синтеза. Возникает все большая
взаимозависимость ассоциативных систем в работе целостной интегративной системы
полушарий, прежде всего в обеспечении процессов кратко— и долговременной памяти
и формировании вероятностных программ поведения на основе доминирующей
мотивации.
3.17.4. Эволюция интегративной деятельности мозга
После оценки уровня
совершенствования высшей нервной деятельности и для создания представления о
путях ее прогрессивного развития нужны соответствующие тесты. С помощью
объективного подхода они должны оценить основные проявления высшей нервной
деятельности: аналитико—синтетические способности мозга и способность к
формированию поведенческих программ. Эти функции мозга зависят от уровня
организации его ассоциативных систем.
Одним из тестов может
служить рекомендованный И. П. Павловым условный рефлекс на комплексный
разномодальный раздражитель, заключающийся, например, в одновременном
предъявлении светового и звукового сигналов. При этом истинный интермодальный
синтез, способный выступать в качестве самостоятельного условного сигнала,
формируется лишь при устойчивом и полном угашении и дифференцировании компонентов,
их комплекса.
Насекомоядные (ежи) и
грызуны (крысы, хомяки) не способны выработать такой условный рефлекс. Каждый
сигнал в отдельности — компоненты и комплекс — сохраняет свое пусковое
значение. Длительная тренировка приводит к невротическим расстройствам
условнорефлекторной деятельности.
Хищные (кошки, собаки) справляются с
аналогичной экспериментальной задачей относительно быстро и с помощью
регулярной тренировки достигают дифференцирования компонентов комплекса.
Следовательно, хищные обладают способностью к интеграции разномодальных
сигналов в целостный образ.
Для приматов (низшие обезьяны)
выработка условных рефлексов на комплекс представляет собой легкую задачу, ибо
в процессе применения комплексного раздражителя компоненты самопроизвольно
утрачивают сигнальное значение и сформированные условнорефлекторные связи
сохраняются месяцами без дополнительной тренировки. Это свидетельствует о более
высоком уровне аналитико—синтетической деятельности обезьян в сравнении с
хищными.
Другой важной стороной интегративной
функции мозга является степень развития процессов памяти и основанное на них свойство
прогнозирования предстоящего поведения. Прогностическая функция мозга до
сих пор мало изучена, и наши знания пока ограничиваются лишь общими схемами.
Одна из них _ это схема функциональной системы поведенческого акта. Согласно
этой схеме, мозг животных способен к формированию не только конкретных
поведенческих программ, но даже физико—химических параметров предстоящего
результата их выполнения. В этой схеме допускается существование только
жесткого программирования. Однако в естественных условиях существования любая
поведенческая адаптация относительна и имеет вероятностный характер. Поэтому применительно
к биологическим условиям обитания животных в постоянно изменчивой внешней среде
имеет место и вероятностное прогнозирование, а значит, адаптивность
поведенческих программ определяется степенью их избыточности и подвижности.
У животных изучали поведение в
стационарных случайных средах: на условный сигнал животное должно было
направляться к одной из двух кормушек, причем чаще всего к той, где наиболее
вероятно получало подкрепление.
Насекомоядные и грызуны совершенно
не способны справиться с такой задачей, их поведенческие программы обладают
низкой лабильностью и характеризуются автоматизмом и стереотипией.
Хищные способны к формированию
предпочтения, той кормушки, вероятность пищевого подкрепления из которой
наиболее высока, причем поведение животных меняется в соответствии с изменением
вероятности подкрепления. Повреждение лобных отделов неокортекса лишает
животное способности к вероятностному прогнозированию.
Приматы легко справляются с задачами
на вероятностное прогнозирование, однако после повреждений в лобной
ассоциативной коре лишаются этой способности. Их поведение приобретает
однообразный стереотипный характер.
Очевидно, способность использования
предыдущего опыта, записанного в долговременной памяти для прогнозирования
поведения в стационарных случайных средах, претерпевает существенные
эволюционные преобразования, которые определяются степенью развития
ассоциативных систем мозга.
3.17.5. Онтогенез ассоциативных систем мозга
Динамика формирования ассоциативных
систем носит гетерохронный характер. Последнее обусловлено включением
отдельных звеньев этих систем s обеспечение разных поведенческих актов, последовательность
созревания которых определяется их необходимостью для выполнения жизненно
важных функций новорожденного животного.
Согласно концепции системогенеза (П. К. Анохин,
1968), неравномерность созревания нервных элементов и связей между ними
объясняется их вовлечением в структуру различных функциональных систем.
Гетерохронность их созревания определяется значимостью для выживания организма
особенно в критические периоды жизни, когда новорожденное животное вступает в
непосредственное соприкосновение с окружающей средой.
Анатомически и функционально первыми созревают те
рефлекторные механизмы, которые обеспечивают выживание на начальных этапах
индивидуального развития. В ассоциативных ядрах таламуса обнаружены их проекции
на кору больших полушарий уже у новорожденных животных. Однако окончательное
созревание ассоциативных систем происходит в течение длительного времени (у
хищных — к 3—му месяцу жизни). В этот период у животных впервые обнаруживают
способность к выработке условного рефлекса на одновременный комплекс.
Затем наступает период бурной дифференциации мозговых
структур и возникновение между ними двусторонних связей диффузного характера.
Формируется та структурно—функциональная матрица, которая служит основой
для дальнейшего развертывания процессов координации и выделения локальных
функциональных структур. Принцип развития — от диффузного неспецифического к
локальному специфическому — общебиологическая закономерность, которой
подчиняется динамика развития ассоциативных систем мозга.
Наконец, последний этап связан с процессом формирования
тормозных координационных механизмов в самой коре и в глубоких структурах.
Появление таких механизмов обеспечивает тонкую специализацию ассоциативных
систем мозга и формирование сложных форм аналитико—синтетической деятельности и
вероятностного прогнозирования.
3.18. ФУНКЦИОНАЛЬНАЯ СТРУКТУРА ПОВЕДЕНЧЕСКОГО АКТА
Наиболее важным звеном в системе адаптивных
свойств организма и динамике эволюционных преобразований поведения является
способность к формированию планов, программ поведения.
Сам по себе условный рефлекс есть не
что иное, как экстраполирование прошлого опыта с учетом настоящей ситуации на
предстоящее будущее. При всем этом процессы и механизмы формирования
поведенческих программ еще окончательно не изучены.
Важно иметь в виду вероятностный
характер любой поведенческой программы. Степень вероятности ее зависит не
только от внешних факторов, но и от внутренней биологической потребности,
преобладающей в данный момент. Моделирование потребного будущего — высшее
отражение активности мозга, который сталкивается с динамически
переменчивой ситуацией, ставящей его перед необходимостью выработки свойств
вероятностного прогнозирования
Программа
поведенческого акта — это модель того, что произойдет с организмом в
будущем; ее можно рассматривать как формирование логики, алгоритма,
функциональной структуры предстоящего поведенческого акта. Такая функциональная
структура опирается на прошлый жизненный опыт, записанный в долговременной
памяти, и актуальное настоящее, куда входит не только изменчивая окружающая
среда, но и сам организм с его потребностями В результате планируется будущий
поведенческий акт, в котором организм должен с той или иной вероятностью
предвидеть возможные изменения в изменчивой среде.
Следовательно,
в основе вероятностного прогнозирования лежит предвосхищение будущего,
основанное на структуре прошлого опыта и информации о наличной ситуации.
Способность к вероятностному прогнозированию есть результат биологической
эволюции в вероятностно организованной среде. С расширением среды обитания и
рецептивных возможностей животного неопределенность окружающей среды возрастает
в динамике эволюции и в индивидуальном развитии, что обеспечивает шансы особи
на выживание.
3.18.1. Основные поведенческие доминанты
Любая поведенческая программа строится на трех
основных детерминантах: доминирующей мотивации, прошлом жизненном опыте (долговременная
память) и оценке текущей ситуации.
Доминирующая мотивация. В основе жизненного опыта лежит
определенная биологическая, а для человека прежде всего социальная потребность
вызвав избирательную активацию определенных мозговых аппаратов она приводит к
созданию доминирующей мотивации. В каждый момент времени доминирующей
становится та биологическая мотивация, в основе которой лежит наиболее важная
потребность. Доминирующая мотивация как первичный системообразующий фактор
определяет все последующие этапы мозговой деятельности по формированию
поведенческих программ. Специфика мотиваций определяет характер и «химический
статус» внутрицентральной интеграции и набор вовлекаемых мозговых аппаратов. В
качестве полезного результата определенного поведенческого акта выступает
удовлетворение потребности т е снижение
уровня мотивации.
Прошлый опыт, долговременная память. Широкий репертуар готовых
функциональных элементов, навыков,
которые либо предопределены генетически, либо сформированы в
индивидуальной жизни, т. е. приобрели
автоматизированный характер, является фундаментом для построения сложных
поведенческих программ.
Поведенческую программу можно рассматривать как систему
интрацентральных условных рефлексов,
представляющих собой комплекс
временных связей между хранящимися в памяти относительно жесткими
сенсомоторными синтезами.
Доминирующая мотивация извлекает из долговременной
памяти готовые элементы, которые в определенных условиях могут оказаться
достаточными для достижения приспособительного эффекта. Последнее возможно при
стабильной неальтернативной окружающей ситуации, которая в данный момент
времени обнаруживает свои жесткие связи и отношения.
Естественный отбор разделил целый
ряд готовых поведенческих реакций на конкретные, регулярно повторяющиеся
жизненные ситуации, которые при возникновении определенной биологической
мотивации реализуются в поведении, не требующем предваряющего программирования
(инстинкты, стереотипные навыки и пр.). Таким путем естественный отбор
обеспечивает организму оптимальные условия встречи с жизненно важными факторами
вероятностно организованной среды. Эта закономерность совершенствуется не
только в динамике филогенеза, но и на ранних этапах индивидуального развития. В
этот период мозг обладает чрезвычайно высокой запечатлеваемостью к биологически
значимым факторам среды. Последнее обеспечивает набор жестких поведенческих
навыков, которые сохраняются в памяти взрослого организма и включаются в
структуру более сложных поведенческих актов.
Оценка текущей ситуации. Для адекватного программирования поведения организму
необходимо: 1) оценить собственную схему тела, вписанного в окружающую среду;
2) извлечь биологически полезную информацию из этой среды;
3) описать структуру среды как закон
связей между ее наиболее существенными переменными; 4) определить ведущее
кинематическое звено для выполнения предстоящего двигательного акта.
Доминирующая мотивация превращает
реальную среду в актуальную. Активная природа процесса восприятия
осуществляется в пределах целостной сенсорной функции мозга и состоит из
операций: обнаружения и кодирования, распознавания и декодирования. Высший
уровень сенсорной интеграции завершается формированием образа среды, оценкой
взаимоотношений между биологически значимыми признаками, т.е. формированием
структуры среды, логики ее построения, конструкции и закономерностей ее
изменений. Животные могут выполнять поведенческий акт в новой обстановке,
оперируя эмпирически усвоенными законами, лежащими в основе структуры среды.
Это свойство, определяющее элементарную рассудочную деятельность животных,
развивается в эволюции и достигает наивысшего совершенства у высших приматов и человека.
Организм стремится к оптимальной
вероятности встречи с биологическими значимыми предметами и явлениями
окружающего мира. Организм извлекает информацию, т. е. упорядоченность для
формирования логики своего поведения. Во всех этих процессах существенная роль
отводится кратковременной памяти, которая необходима для удержания
следового образа окружающей среды и программы предстоящего поведенческого акта
вплоть до его полной реализации.
3.18.2. Ассоциативные системы мозга
и структура поведения
Систему операций, лежащих в основе
функциональной структуры поведенческого акта, невозможно локализовать в
определенных анатомических субстратах мозга. Наоборот, важно оценить степень
участия различных мозговых аппаратов в целостной мозговой функции.
Существует ряд условий, на которых ту или иную мозговую
систему можно рассматривать как полноправного участника программирующей функции
мозга. Эти условия следующие. Мозговая система должна получить информацию
|
|
|
Рис. 3.55 Функциональная
структура , целенаправленного
поведенческого акта Сплошными
стрелками обозначены взаимоотношения между блоками, пунктирной — пути
безусловных и условных автоматизированных рефлексов. |
о доминирующей в данный
момент мотивации, о репертуаре навыков в долгосрочной памяти, о факторах
внешней среды и схеме тела. Кроме того, мозговая система должна обладать
внутренними свойствами к фиксации следовых процессов в кратковременной памяти и
организации селективного внимания. Наконец, эти системы должны обладать
предпосылками для координированного управления остальными аппаратами мозга.
Этим условиям
отвечают ассоциативные таламокортикальные системы мозга. Таламопариетальная
система на основе доминирующей мотивации обеспечивает селективное внимание к
сигнально значимым факторам окружающего пространства и ведущего кинематического
звена схемы тела, а также участвует в механизмах управления позой.
Таламофронтальная система обладает механизмами для ее участия в вероятностном
прогнозировании и завершает организацию программы целенаправленного действия
согласно доминирующей мотивации, прошлому жизненному опыту и наличной
окружающей ситуации.
Вероятностные
программы формируют команды, т. е. актуализируют в строго временной зависимости
набор поведенческих тактик (блоков), в результате чего совершается действие,
которое может привести к получению или отсутствию результата (рис. 3.55).
Достижение последнего результата сопровождается переводом данной программы в
долгосрочную память, а эффективность действия оценивается по степени
выраженности положительных эмоций.
Система эмоций —
единственная организация, способная осуществлять оценку степени соответствия
действий организма доминирующей мотивации и прогнозируемой вероятности ее удовлетворения.
При отсутствии ожидаемого результата включаются различные
механизмы: 1) сохранения программы в кратковременной памяти и повторение того
же действия; 2) отрицательных эмоций, сигнализирующих о неуспехе действия, что
может привести к изменению самой мотивации; 3) стойкой мотивации, побуждающей к
перестройке самой программы действия, т. е. выдвижению новых гипотез и
формированию новой функциональной структуры. Если организм находится в
стационарной среде, динамика изменений которой стабильна и регулярно
повторяется, то можно допустить существование более простых мозговых механизмов
типа безусловных рефлексов или автоматизированных условных рефлексов, которые
выполняются системой жестких внутримозговых связей.
Адаптивное поведение в постоянно меняющейся случайной среде
требует выработки способностей к вероятностному прогнозированию и
соответствующих пластических мозговых систем,
коими и
являются эволюционно молодые ассоциативные системы мозга.
3.19. ОСОБЕННОСТИ ВЫСШЕЙ НЕРВНОЙ (ПСИХИЧЕСКОЙ) ДЕЯТЕЛЬНОСТИ
ЧЕЛОВЕКА
При изучении психической
деятельности человека И. П. Павлов большое значение придавал пониманию ее
физиологических основ и закономерностей. Предложенный им объективный метод
условных рефлексов рассматривали как важнейший инструмент, с помощью которого
создавался фундамент психологического знания.
3.19.1. Физиологические основы
психики
Психические процессы теснейшим
образом связаны с физиологическими явлениями, но не сводимы к ним. В основе
психических явлений лежат не элементарные процессы возбуждения и торможения, а
системные процессы, объединяющие отдельные физиологические проявления в
интегрированное целое. Принцип доминанты А. А. Ухтомского объясняет, как из
множества отдельных рефлекторных актов складывается единство.
Понятие об интегральном образе
как продукте пережитых доминант, т. е. как субъективном отражении
объективного взаимоотношения организма со средой, сыграло принципиальную роль в
решении проблемы соотношения психического и физиологического.
Важный шаг в ее развитии был сделан
П. К. Анохиным в его концепции функциональных систем, где система, в
которой участвуют физиологические процессы, является внутренним отражением
внешней среды. Системные процессы поведенческого акта — это информационные
процессы, для которых физиологические процессы выступают как материальный
носитель. В этом смысле психический образ и его нейродинамический эквивалент не
только одновременные явления, но они составляют нераздельную целостность.
Теория отражения представляет
психику как свойство высокоорганизованной материи отражать своими состояниями
независимо от них существующую действительность. Психическое закономерно
возникает на определенной ступени развития жизни, т. е. усложняющиеся условия
ставят организм перед необходимостью отражения объективной действительности в
виде ощущений и восприятии, ведущих к ее осознанию. Противоположность между
субъективным и объективным не изначальна, не абсолютна, она порождается
развитием определенных форм отражения.
Учение о доминанте А. А. Ухтомского
позволяет ответить на основной вопрос: что превращает сумму отдельных реакций в
целенаправленные поведенческие акты с новыми, ранее неведомыми свойствами? Это
один из нейрофизиологических механизмов психической деятельности — не только
внешней, предметно развернутой, но и внутренней. Психические состояния и
процессы «опредмечиваются» как интегральные образы, в основе которых лежат
корковые доминанты. Последние то активно действуют в поле сознания, то
переходят в сферу неосознанного, где сохраняются в скрытой форме как следы прошлой
деятельности. Такие доминанты могут либо поочередно «выплывать» в поле ясного
внимания, либо вступать в конфликт, и тогда душевная жизнь будет борьбой
вытесняющих друг друга «несогласных» доминант.
Всякое понятие и представление,
всякое индивидуальное психическое содержание является отражением пережитой
некогда доминанты. Этот след, а иногда и вся доминанта в полном ее выражении
вплоть до эмоциональных состояний и вегетативных сдвигов могут воспроизводиться
адекватным сигналом. Указанные явления лежат в основе механизмов воспоминания и
мышления.
|
|
|
Рис.
3.56 Сопоставление сложнейших безусловных рефлексов (инстинктов) высших
животных с потребностями человека |
А. А. Ухтомский боролся с биологизацией социальных проблем и
указы вал, что понятие «среда» содержит в себе некий общественный момент— род
вид, общество и индивид должны сообразовывать свою деятельность с интереса' ми
этого общества. Человек — продукт общества, и сущность его поведения должна
быть понята как результат сочетания накопленного опыта поколений знании,
традиций, усвоенных данным индивидом, и его личных способностей и особенностей.
С одной стороны, сознание осуществляет свою деятельность на основе корковых
доминант, с другой — оно само стремится воспитать такие доминанты, которые, бы
содействовали успешному преобразованию характера и направленности
действий.
Это означает, что даже
механизмы гомеостаза не могут быть полностью лишь физиологическими. Гомеостаз у
человека вышел за пределы границ индивида и стал определять отношения между
людьми. Поэтому механизм гомеостаза полностью не раскрывает направленность
реакций организма на сближение с раздражителями внешней среды, на его
приспособление к действующим факторам среды. Доминанта как нарушение равновесия
между центрами, с ее выраженной экспансией и нацеленностью, ведет к обогащению
организма новыми возможностями.
Поэтому предложена классификация
человеческих потребностей трех основных, независимых по происхождению
групп: витальных, социальных и идеальных потребностей познания и
творчества. В каждой из этих групп выделяют потребности сохранения и развития,
а в группе социальных — потребности «для себя» (права) и «для других» (обязанности}.
Любая из перечисленных выше потребностей может быть удовлетворена при наличии
дополнительных потребностей: вооруженности средствами, знаниями, умениями в
преодолении препятствий на пути к цели, обычно именуемые волей (рис.
3.56).
Сфера потребностей человека является
основой для создания информационной сферы, ядром которой выступает сознание.
3.19.2. Сознание и неосознаваемое
Специфически человеческая форма
отражения действительности представлена сознанием, т. е. таким знанием,
которое с помощью слов, математических символов, образов, художественных
произведений может быть передано другим людям и другим поколениям в виде
памятников культуры.
В самостоятельную группу явлений
выделяют неосознаваемые психические явления, куда входит все то, что либо
было познанным, либо при определенных условиях может стать осознанным. В мозгу
допускают существование чувствительного механизма, реагирующего на очень
слабые, но психологически весьма значительные для данной личности раздражители.
Сам этот механизм не обеспечивает осознания эмоционально значимого
раздражителя, но его активация приводит к электрическим, вегетативным и
психическим сдвигам. Такой механизм получил название психологической защиты.
3.20. ФУНКЦИОНАЛЬНАЯ МЕЖПОЛУШАРНАЯ
АСИММЕТРИЯ
После открытия в левом полушарии моторного
центра речи (центр Брока) и сенсорного центра речи (центр Вернике)
это полушарие стали рассматривать доминирующим в отношении речевой функции и
мышления. Под влиянием наблюдений за больными людьми с перерезкой спаек
(комиссур) переднего мозга («расщепленный мозг») стала складываться концепция о
частичном доминировании полушарий мозга человека. Согласно этой
концепции, левое полушарие специализируется на вербально—символических
функциях, правое — на пространственно—синтетических. Различия между
полушариями при зрительном восприятии показаны в табл. 3.4.
Из анализа приведенных данных следует существование пяти
основных различий полушарного доминирования у человека: вербально—невербальное,
время—пространство, анализ—синтез, последовательное—одновременное восприятие,
абстрактное и конкретное узнавание.
Та б л и ц а 3.4
Межполушарные различия при зрительном восприятии
|
Левое
полушарие |
Правое
полушарие |
Лучше узнаются
стимулы
|
|
Вербальные
|
Невербальные
|
|
Легко
различимые |
Трудно
различимые |
|
Знакомые |
Незнакомые
|
Лучше решаются
задачи
|
|
|
Оценка
временных отношений |
Оценка
пространственных отношений |
|
Установление
сходства |
Установление
различий |
|
Установление
идентичности стимулов по названиям |
Установление
физической идентичности стимулов |
|
Переход к
вербальному кодированию |
Зрительно—пространственный
анализ |
Особенности
процессов восприятия
|
|
|
Аналитическое
восприятие |
Целостное восприятие (гештальт) |
|
Последовательное
восприятие |
Одновременное
восприятие |
|
Абстрактное,
обобщенное, инвариантное
узнавание
|
Конкретное
узнавание |
Предполагаемые
морфофизиологические различия
|
|
|
Фокусированное
представительство |
Диффузное
представительство |
Выделяют несколько видов функциональных асимметрий.
Неодинаковость двигательной активности рук. ног, лица, половин тела,
управляемой каждым полушарием мозга, называется моторной асимметрией.
Неравнозначность восприятия каждым из полушарий объектов, расположенных слева и
справа от средней плоскости тела, именуется сенсорной асимметрией.
Специализация полушарии мозга в отношении различных форм психической
деятельности обозначается как психическая асимметрия.
Так, человек с превалированием
левополушарных функций тяготеет к теории, имеет большой словарный запас и
активно им пользуется, ему присуща двигательная активность, целеустремленность,
способность прогнозировать события. Правополушарный» человек тяготеет к
конкретным видам деятельности, он медлителен и неразговорчив, наделен
способностью тонко чувствовать и переживать, он склонен к созерцательности и
воспоминаниям.
Совокупность индивидуальных особенностей психики и поведения
человека составляет тип высшей нервной деятельности, или темперамент человека
ин складывается из общих свойств нервной системы, которые характеризуются
экстра—интраверсией, эмоциональной
стабильностью — невротизмом, и подвижностью или инертностью нервных процессов.
И. П. Павлов выделял два типа высшей нервной деятельности человека,
которые сейчас могут рассматриваться с позиции межполушарной
асимметрии—мыслительный тип с преобладанием второй (речевой) сигнальной системы
—преимущественно левополушарный и художественный тип с преобладанием первой
сигнальной системы конкретных образов — преимущественно правополушарный
человек. В последнее время получает признание концепция о взаимодополняющем
сотрудничестве двух полушарий и преимуществе отдельного полушария лишь в
определенные стадии той или иной нервно—психической деятельности, а не всей
функции в целом. Правое полушарие быстрее, чем левое, обрабатывает поступающую
информацию, зрительно—пространственный анализ стимулов в правом полушарии
передается в левое полушарие (в моторный «центр речи»), где происходит
окончательный, высший семантический анализ и осознание раздражения.
Согласно концепции В. Л. Бианки, в процессе обучения правое
полушарие работает по принципу дедукции, т. е. сначала осуществляет
синтез, а затем анализ; левое же полушарие функционирует по принципу индукции,
сначала анализируя раздражители, а затем синтезируя их (рис. 3.57).
В эволюции человека речь первоначально лишь использовала, а
затем развила и качественно преобразовала уже имеющуюся у животных
функциональную межполушарную асимметрию.
|
|
|
Рис. 3.57 Схема межполушарных взаимоотношений (по Бианки) Вверху — ассоциативная кора, внизу
— проекционная; слева — левое полушарие; справа — правое; жирные стрелки —
доминирующие влияния, тонкие — недоминирующие; белые стрелки — облегчающие
влияния; прерывистые — тормозящие; 1,5
— транскаллозальные (через мозолистое тело) влияния; 2,7, 10 — восходящие афферентные
влияния; 3, 8 — дивергенция
возбуждения; 4 —
конвергенция; 6 —
экстракаллозальные влияния; 9 —
межзональные транскаллозальные влияния; 11 — межзональные внутриполушарные влияния; 12, 13 — транскаллозальные
облегчающие влияния; 14, 15 —
транскаллозальные тормозящие влияния; 16
— экстракаллозальные облегчающие влияния; 17 — экстракаллозальные тормозящие влияния. |
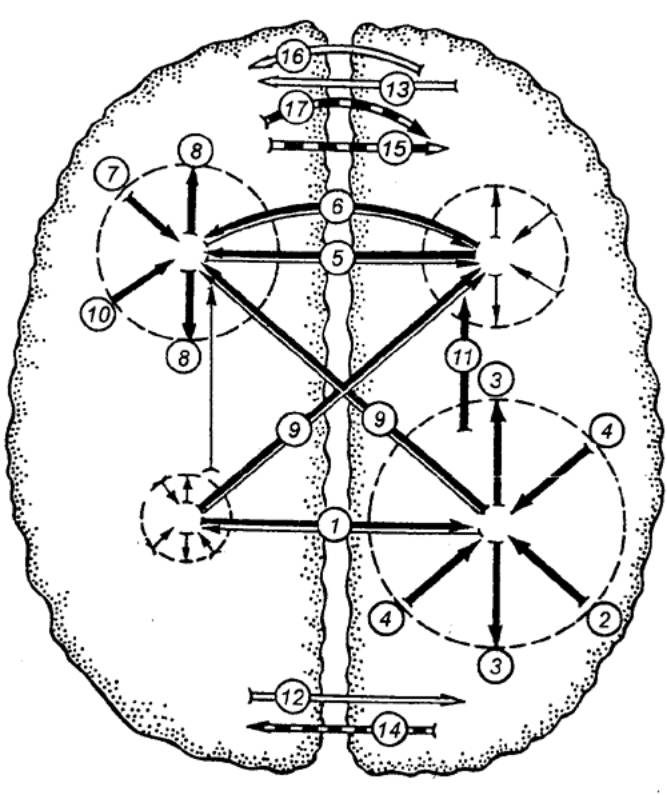
3.21. ФОРМИРОВАНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ РЕБЕНКА
Современные представления о нервных
механизмах поведенческих и психических процессов строятся на признании определяющей
роли ассоциативных систем мозга. Эти образования мозга участвуют в
выполнении многих высших корковых функций: программировании целенаправленных
движений и действий, многих интеллектуальных операциях, познавательных
процессах, организации системы речевого высказывания, наглядных
пространственных и символических синтезов, формировании кратковременной
пространственной и речевой памяти.
Формирование высшей нервной деятельности ребенка тесно
связано с динамикой созревания ассоциативных корковых структур. Ребенок в
общении со взрослыми усваивает определенные знания, приобретает новые формы
отношений к внешнему миру. Все это строится на основе создания новых
функциональных связей, которые позволяют овладеть новыми формами восприятия и
запоминания, видами мышления, способами организации движения. Их структурной
основой выступают лобные и нижнетеменные области коры, которые в
онтогенезе человека увеличиваются по площади в 9 раз и у взрослого занимают
|
|
|
Рис. 3.58 Топографическое
расположение моторного (зона Брока, I), слухового (зона Вернике, II) и
зрительного (III) центров речи левого полушария мозга человека |
|
|
|
Рис. 3.59 Локализация
функций в коре большого мозга человека |
около 2/3 всей поверхности новой
коры. Кроме того, у человека в онтогенезе развиваются специфические
человеческие корковые формации, которые имеют специальное отношение к анализу и
синтезу раздражении, связанных с разными сторонами речевой деятельности (речедвигательной,
речеслуховой, речезрительной) (рис. 3.58, 3.59).
Установлено, что в течение первого года жизни ребенка
количество условных рефлексов на непосредственные раздражители постепенно
увеличивается. Вначале формируются только вегетативные условные рефлексы, затем
двигательные и, наконец, появляются речедвигательные. Наличие последних не
свидетельствует еще о появлении словесного мышления, так как нет еще отвлечения
и обобщения при помощи слов.
Исходно слово выступает как слабый компонент комплексного
раздражителя и имеет второстепенное значение. Однако постепенно при условии
частого повторения слово как наиболее постоянный компонент комплекса
приобретает значение сильного компонента и, наконец, самостоятельного
сигнала, хотя к концу 1—го года жизни ребенка первые воспринимаемые им
слова — это просто
.
|
|
|
Рис. 3.60 Формирование механизмов восприятия слова у детей в процессе развития А —
новорожденные; Б—в возрасте 3—10 месяцев. В—в возрасте 2—3 лет: а — уровень рецептивных
преобразований, б — уровень
фонемного перекодирования слова, в — уровень семантического анализа слова, г — зоны мозга, участвующие в
семантическом анализе слова; 1 —
лобные зоны, 2 — моторные, 3
— нижнетеменные, 4 —
затылочные, 5 — височные; Заштрихованной стрелкой обозначено воздействие
слова. |
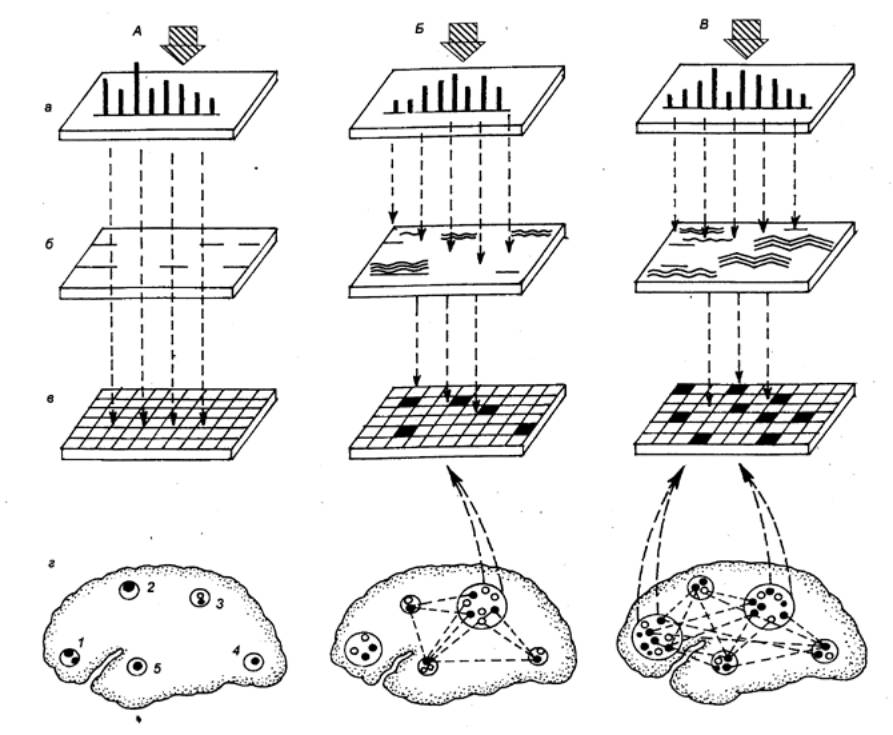
звуковые обозначения определенных
предметов и лиц. Позже, в следующем возрастном периоде, происходит превращение
слова в абстрактный сигнал, обозначающий совокупность предметов.
На 2—м году жизни возникают образы отдельных предметов.
В результате действия ребенка с предметами они вычленяются из обобщенного
недифференцированного мира. Формирование речевых реакций невозможно без
звукового словесного подражания. При этом особую роль играет установление связи
(ассоциации) слова с видами предмета, его осязанием и другими признаками.
Различают понимание речи (сенсорная речь) и ее воспроизведение (моторная речь).
Вначале возникает понимание речи, затем оба эти процесса теснейшим образом
увязываются.
Развитие обобщающей функции слова
проходит через следующие основные этапы.
I степень интеграции — слово
замещает чувственный образ определенного предмета, т. е. оно является
эквивалентом одного конкретного предмета («ляля» — только данная кукла; конец
1—го — начало 2—го года).
II степень интеграции — слово
замещает несколько чувственных образов однородных предметов («ляля» относится к
нескольким куклам; конец 2—го года).
Ш степень интеграции — слово
замещает ряд чувственных образов разнородных предметов («игрушка» — это и
куклы, и мячи, и кубики; не раньше 3—го года).
IV степень интеграции — в слове
связан ряд обобщений предыдущих степеней (например, слово «вещь» содержит
обобщение, даваемое словом «игрушка», «одежда», «еда» и др.; 5—й год жизни).
С помощью электроэнцефалографических
исследований установлена динамика функционального созревания основных корковых
зон у детей. Ведущим звеном в организации межцентральных отношений выступают
ассоциативные отделы неокортекса — нижнетеменные и лобные зоны (рис. 3.60). У
детей 1—го года жизни фокусами взаимосвязной активности являются нижнетеменные
зоны, которые устанавливают связи с затылочными, височными и моторными центрами
коры. У 2—летних детей фокусы взаимосвязной активности перемещаются в лобные
отделы, которые вступают в синхронные взаимоотношения с другими центрами коры.
У детей 4—5 лет отмечается усиление межцентральных взаимодействий нижнетеменных
полей. На более поздних этапах онтогенеза (6—7 лет) вновь происходит усиление
межцентральных связей лобных областей с другими отделами коры.
V годовалых детей
только при опознании знакомого слова усиливаются связи нижнетеменных и височных
областей коры левого полушария. На 2—м году жизни при опознании знакомого слова
усиливаются связи двух ассоциативных областей — нижнетеменных и лобных
преимущественно левого полушария.
У детей 4—5 лет при восприятии
знакомого слова связи левой лобной и левой нижнетеменной зон еще более
усложняются. Такая динамика отражает морфофункциональную зрелость высших
интегративных систем мозга ребенка.
Начиная с 6—летнего возраста ребенок
в состоянии управлять своим поведением на основе предварительной
словесно—обобщенной инструкции. К этому возрасту он способен удерживать
программу действий, состоящих из ряда движений. Поведение ребенка начинает в
большей степени определяться степенью вероятности тех или иных событий в
окружающей среде. К этому возрасту более ярко проявляются типологические
особенности высшей нервной деятельности (по признакам силы, уравновешенности и
подвижности процессов возбуждения и торможения). К 10—11—летнему возрасту
развитие коры головного мозга приближается к уровню взрослого человека, что
является важнейшим фактором формирования высших нервных и психических функций
ребенка.
Мышление — сложнейший вид мозговой
деятельности человека в процессе приспособления к новым условиям и при решении
новых жизненных задач. Процессы мышления сводятся к образованию общих
представлений и понятий, а также суждений и умозаключений. Помимо
словесно—логического, абстрактного существуют формы эмоционального мышления
(оценка), практического, или наглядно—действенного, мышления.
Среди множества теорий, пытающихся объяснить
природу мышления, наиболее распространены две: одна рассматривает мышление
как беззвучную речь, как выражение работы речевого механизма (и механизма
письма), а у глухонемого — механизма жестикуляции; вторая — считает мышление
своеобразным чисто психическим процессом, лишенным образности и не совпадающим
с внутренней речью. Полного подтверждения не получила ни одна из этих гипотез.
Раскрытие действительной сложности обоих процессов:
превращения мысли в речь и извлечения основного содержания (мысли) и речевого
сообщения — было связано с успехами психологии и лингвистики.
Мысль является сложнейшим обособленным отражением
действительности, направляющимся известными мотивами, т. е. это специфический
процесс, сформированный вещественно—историческим развитием на базе той роли,
которую играет язык в истории человечества. Поэтому мысль, которая на начальных
этапах истории сама являлась конкретной развернутой деятельностью и которая
лишь со временем превратилась в свернутый внутренний процесс, не может
рассматриваться как изначальный «духовный» акт.
В онтогенетических исследованиях установлено, что на ранних
этапах развития ребенка всякое препятствие, формирующее «задачу», вызывает
возникновение внешних проблем, в которые включается анализирующая и планирующая
внешняя речь. На последующих этапах развития эта развернутая внешняя речь,
включающаяся в действие и способствующая решению задачи, переходит в шепотную
речь, и именно эта внутренняя речь, еще более концентрируясь, становится
основой глубинного процесса, который на конечных стадиях превращается в мысль.
Восприятие речи является важнейшей стороной функционирования
мозга человека. Обработка речевого сигнала проходит по законам сенсорных
систем, хотя обладает специфическими особенностями, определяемыми сложной
иерархической структурой речи. Речь как сигнал имеет два класса признаков:
собственно физические признаки (фонемы, слоги) и информационные (просодические)
признаки, состоящие в интегрировании отдельных сегментов в более крупные
единицы — слова, фразы.
В физиологии речевой функции обнаруживаются основные
признаки отражения внутреннего (субъективного) и внешнего (объективного) мира.
Это должно включать в себя прежде всего механизм выделения существенных
признаков внешних стимулов и отношения этих признаков, которые формируют основу
для обобщения. Другой составной частью механизмов речевой функции является
обеспечение единства целенаправленной деятельности, сохранение задачи и
интеграции замысла, лежащих в основе внутренней свернутой речи, которая
является базой для реализации развернутой (экспрессивной) речи. Функция речи
выполняется при обязательном взаимодействии этих двух механизмов: гностического
и динамического.
Локальные поражения левого полушария у правшей приводят к
нарушению речевой функции в целом, а не к частичному выпадению какой—либо одной
ее формы. Распад экспрессивной речи, связанный с нарушением динамических схем
слов, неизбежно вызывает распад единства звука и смысла и, как следствие этого,
нарушение понимания речи.
Клинические исследования механизмов речепродукции и
речевосприятия указывают на существенную роль кортикальных формаций прежде
всего левого полушария и подкорковых образований — различных структур переднего
и промежуточного мозга. Существует предположение о конвергенции механизмов,
обеспечивающих раздельное восприятие и продукцию речи, которая усиливается от
нижних этажей мозга к высшим, достигая максимума на элементах лобного полюса
коры.
|
|
|
Рис.
3.61 Электроэнцефалографические
стадии сна, зарегистрированные у ребенка 12 лет А—Е — обозначения соответствующих стадий по классификации
Лумиса, Р — парадоксальный
сон. |
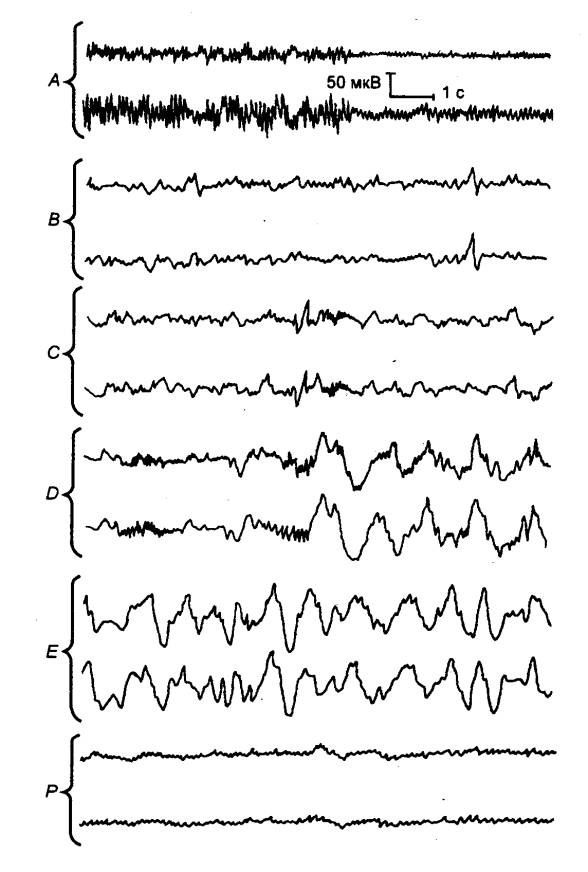
Для спокойного бодрствования
и дремоты (рис. 3.61) характерен α—ритм, амплитуда которого постепенно
уменьшается и может вообще исчезать (стадия А). При внешнем раздражении
наступает реакция активации, т. е. происходит смена α—ритма на
высокочастотный (β—ритм (реакция десинхронизации). При появлении легкой
сонливости (стадия В) возникает неустойчивая высокоамплитудная
активность. Стадию С связывают с поверхностным сном, когда начинают
появляться низкокоамплитудные медленные волны. Переход к стадии D и особенно Е свидетельствует
о глубоком сне, который сопровождается дальнейшим усилением
высокоамплитудной медленной активности (S—ритм). Типичная картина
парадоксального сна (ПС) — низкоамплитудная нерегулярная активность с
непостоянными редкими группами α—ритма и вспышками острых пилообразных волн.
Общая картина сна детей существенно
меняется с возрастом: если новорожденный бодрствует около 4 ч/сут, то к концу
первого года жизни время бодрствования возрастает до 10 ч/сут. У взрослых в
течение ночи отмечается 4—6 циклов ПС длительностью каждый по 20 мин. Если у
новорожденных общая продолжительность быстрого сна занимает около 10 ч, то у
взрослых — только 1,5 ч. Однако значение ПС для обеспечения продуктивного
бодрствования с возрастом увеличивается. Продолжительность сна взрослых людей
очень индивидуальна.
Сновидения. Сновидениями интересовались испокон
веков. Их трактовка дается еще в ведических текстах древней Индии. Мощнейший
импульс к экспериментальному изучению сновидений дало открытие ПС. Было
установлено:
1) каждый человек видит сны много
раз в течение ночи; 2) насыщенность сновидения событиями примерно соответствует
реальной длительности сна; 3) испытуемые отчитываются о сновидениях в 80%
случаев при пробуждении после ПС и лишь несколько реже — при пробуждении во
время МВС; 4) во время ПС доминируют яркие, сложные, странные, эмоционально
окрашенные и длительные сновидения, тогда как во время МВС — кратковременные
рациональные и реалистические сны, более сходные с мышлением в бодрствовании;
5) переживание сновидений возникает также при засыпании и при пробуждении,
ночные и дневные грезы близки и по характеру, и по содержанию; 6) при
пробуждении из глубокого МВС отмечаются сонливость, дезориентация и нарушение
памяти, тогда как при пробуждении из ПС — быстрая ориентация и готовность к
действию; 7) более одной трети сновидений содержат эмоции страха и тревоги;
поражения, неудачи и конфликты случаются в сновидениях чаще, чем
удовлетворение, успех и дружеские контакты; странности, фантастика и обыденные
ежедневные дела встречаются нечасто; большинство сновидений банальны и
неинтересны; 8) в сновидениях отсутствуют элементы воображения и трезвая оценка
невероятности развертывающихся событий; 9) сновидения быстро забываются, кроме
того, сновидения имеют особую атмосферу, которую трудно передать словами.
Гипотез о природе сновидений
множество. 3. Фрейд сравнивал сновидения с «работой сна» и рассматривал их как
особый важный язык мозга. Он был твердо убежден, что «толкование сновидений —
это основной путь к познанию бессознательного». Во многих гипотезах
подчеркивается, что состояние сновидений обусловливается в основном активностью
самого мозга. Необыкновенный, пестрый характер сновидений может быть
результатом синтеза мозгом противоречивой информации при одновременной
активации различных нейронных сетей. Ряд авторов рассматривает сновидения как
процесс пересмотра данных, хранящихся в памяти. Молекулярные же биологи
полагают, что сновидения способствуют «очистке» памяти от ненужной информации и
тем самым поддерживают эффективность работы нервной системы. Эти и многие
другие гипотезы трудно подтвердить и опровергнуть. Окончательный ответ о
биологическом значении сновидений еще не найден.
Проблема сновидений — наиболее
загадочная область человеческой психики. Сны преимущественно носят зрительный
характер. У слепорожденных в снах отсутствуют зрительные образы и преобладают
осязательные. На характер сновидений оказывает влияние профессиональная
деятельность человека: музыканты чаще переживают чисто слуховые сновидения.
Отмечается зависимость сновидений от уровня интеллектуальности, пола, возраста,
утомления.
В целом сновидения отражают
имеющиеся у личности проблемы, а материалом является конкретный опыт,
накопленный в течение жизни. И. М. Сеченов называл сновидения «небывалыми
комбинациями бывалых впечатлений». Установлено, что нет людей, не видящих сны.
Сновидения возникают в среднем 4—6 раз за ночь. Если пробуждение наступает в
стадии быстрого сна, 70—90% людей детально и достаточно эмоционально
рассказывает о своих сновидениях, а если в медленном — лишь 7—10%. При этом
психические процессы медленного сна скорее выступают как мышление, пересказ
событий прошедшего дня.
Сновидения иногда рассматривают как
следствие растормаживания нервных следовых процессов, связанных с прошлым
жизненным опытом и социально обусловленных по своему содержанию. Для понимания
механизмов извлечения из памяти и направленного характера сновидений может быть
использована теория доминанты. Скрытые корковые или подкорковые доминанты могут
выступать в качестве основных факторов сновидений, а сами последние могут
возникать со всеми признаками доминанты (стереотипные навязчивые сновидения).
В. М. Бехтерев при анализе
диагностического значения сновидений исходил из следующей позиции: в
сновидениях происходят явления, которые развиваются из того же источника, что и
внутренние переживания в состоянии бодрствования. Сновидения указывают на
наличие в прошлом или в настоящем определенных физиологических или психических
состояний.
Гипноз и внушение. Гипнотическое состояние является
сноподобным состоянием с сохранением речевой связи (раппорта) гипнотика с
гипнотизером. Различают три стадии гипноза: 1) сонливость, при которой
гипнотик может противостоять словесному внушению и открыть глаза; 2) гипотаксия
легкий сон, когда он не может открыть глаза и подчиняется внушению; 3)
сомнамбулизм — глубокий сон, когда гипнотик полностью подчинен гипнотизеру, у
него развивается амнезия (забывание предыдущих событий) после пробуждения.
Гипнабельность, т. е. подверженность гипнозу, степень
и вид внушаемости, зависит от возраста, пола, интеллекта, здоровья, т. е. от
многих индивидуальных качеств личности. Внушение под гипнозом — лишь пример из
общего учения о внушении, которое свойственно каждому человеку. При внушении
происходит вторжение в сознание посторонней идеи или приказа без участия воли и
внимания реципиента на подсознательном уровне его психики. Этим последним
внушение отличается от убеждения, которое формируется с участием сознания. В
отличие от внушения извне существует еще самовнушение, при котором в сознание
внедряется не только определенная мысль, но и эмоциональное ее переживание.
Чистого самовнушения без воздействия на личность извне не бывает, поэтому
источником процесса самовнушения всегда является внушение, т. е. внешние
факторы. Механизм гипноза остается неизвестным, а завершенной теории гипноза
еще не разработано, хотя целый ряд заболеваний успешно излечивают под гипнозом.
Гипноз иногда рассматривают как специфическую разновидность
бодрствования, полагая, что внушенные гипнотический сон и какие—либо действия —
явления одного порядка. Действительно, ЭЭГ человека, находящегося под гипнозом,
сходна с ЭЭГ человека в состоянии бодрствования; гипнотическое внушение
подчиняется закономерностям условнорефлекторной деятельности; у ряда лиц в
состоянии бодрствования можно получить все эффекты гипнотического внушения.
В психологическом плане в основе
гипноза лежит моральная капитуляция человека, который некритически подчиняется
приказу гипнотизера. Действительно, снижение волевых свойств личности,
повышенная тревожность усиливают внушаемость. Во время гипноза высшая нервная
деятельность существенно меняется. Это касается сенсорной сферы, когда можно
внушить отсутствие слуха, зрения, осязания и даже боли, можно получить извращенные
реакции (иллюзии), вызвать галлюцинации (восприятие несуществующих предметов и
явлений или исключить из восприятия определенные предметы). Гипнотику можно
внушить другой его возраст, различные эмоциональные состояния, воздействовать
на чувства, сферу его внимания, но нельзя изменить личность человека вопреки
его социальным и индивидуальным убеждениям.
3.24. ТРУДОВАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА—ОПЕРАТОРА
Современное производство оснащено
значительными техническими средствами, и на человека—оператора, входящего составной
частью в эту систему, ложатся задачи по контролю, управлению и регулированию
целыми комплексами. В то же время он освобождается от физических нагрузок, а
доля умственного труда существенно увеличивается. Оператор в такой системе
становится интегральным звеном управления, поэтому его характеристики
предопределяют успешность и надежность функционирования данной системы.
Ярким примером является водитель
автотранспорта, который воспринимает показания различных приборов в кабине,
сигналы от дороги, внешнего окружения, пешеходов. Он также воспринимает сигналы
от мышц собственного тела, шум двигателя и др. Основываясь на анализе этих
сигналов, разделяя их на значимые и незначимые, он принимает решение
относительно
|
|
|
Рис. 3.62 Система человек—машина,
работающая по замкнутому и открытому контурам управления |
воздействия на органы управления
автомашиной. Последняя вновь подает информацию о новом режиме движений —
образуется так называемая замкнутая система человек—машина (рис. 3.62).
Такие системы составляют большинство и в других областях человеческой
деятельности.
Следовательно, задача
человека—оператора состоит в переработке за короткое время потока различного
вида информации и быстром принятии единственно правильного решения. Такая
ответственность повышает эмоционально—психическую напряженность работающего
человека, а также социально—экономическую значимость его труда для общества.
Обработка информации
человеком—оператором.
Если человека сравнить с компьютером, то у него больше недостатков, чем
преимуществ. Но достоинства человека оказываются решающими, особенно его
способность выполнять задачи логико—индивидуальными методами, что не требует в
каждом случае трудоемкого предварительного программирования. Практический опыт
и полученное образование уже предполагают наличие таких программ. Человек легко
ориентируется в непредвиденных ситуациях, мгновенно восстанавливает в памяти
предыдущие факты и методы решения проблем, ему не нужно специальное кодирование
информации и трансформации ее в числовую форму.
Вместе с этим человек имеет ряд
существенных ограничений, создающих значительные проблемы в его работе. Сюда
относится и ограниченность ресурсов памяти, и медленная скорость обработки
информации, и, что самое главное, подверженность влиянию утомления, измененного
функционального состояния, настроения и других психофизиологических факторов,
которые ухудшают его способность к оценке информации и быстрому принятию
правильного решения.
Диктуемая практикой необходимость
отражать на дисплее все больший объем информации вынуждает изыскивать
возможности расширения ассортимента размерностей и повышения плотности
размещения кодовых знаков. Легче всего кодируется цвет, затем форма, яркость,
размер. Звуковые сигналы оказываются наилучшими для привлечения внимания к
мгновенной опасности. Слуховая информация лучше передает простое, короткое
сообщение, требующее немедленного действия. Информация о последствиях тех или
иных действий оператора образует обратную связь, обеспечивающую саморегулирование
системы.
Рабочие движения человека—оператора. Рабочие движения являются
компонентами деятельности человека и включаются в структуру этой деятельности.
Физиологически они обеспечиваются тремя факторами: динамической формой
мышечной деятельности (собственно движение), статической мышечной работой
(поддержание рабочей позы) и координацией элементов статики и динамики в
необходимых соотношениях.
Эффективность деятельности оператора зависит больше от
совпадения характеристик сочетания сигнал—ответ, чем от специфических черт
отдельно взятых приборов и ответных движений. Сочетание сигнал—ответ должно
основываться на фундаментальных привычках, т. е. полностью согласовываться с
личными и групповыми стереотипами. Рациональная организация рабочих движений
создает условия для снижения утомления и повышения работоемкости, но вместе с
этим требует соответствующей конструкции органов управления.
Временная структура
деятельности оператора. Временная структура деятельности может быть либо свободной,
ориентированной на точностные, качественные показатели, либо навязанной,
когда главной становится скорость выполнения операции. Как правило, оба эти
показателя присутствуют совместно, неразрывно, требуя точности реагирования и
быстроты выполнения. Каждому оператору приходится работать в условиях
определенной степени риска, что вызывает эмоциональную напряженность, резко
возрастающую в аварийной ситуации. В этом случае временной фактор является
решающим, а правильное и отчетливое ориентирование во временном режиме имеет не
меньшее значение, чем точность выбора реакции. Трудовая деятельность строго
нормирована по объему труда и длительности ее осуществления. При разработке
режимов труда и отдыха учитывают периодические колебания работоспособности
человека. Особое значение это положение приобретает для организации сменного и
ночного труда, где суточная периодика работоспособности становится важным
фактором деятельности человека.
Индивидуальные
особенности человека—оператора. Эмоциональное напряжение, вызываемое условиями труда и
сложностью работы, а также сильная эмоциональная реакция, вызванная, например,
аварийной ситуацией, по—разному влияют на оператора. Адекватность его поведения
во многих отношениях будет определяться индивидуальными особенностями. Состояние
эмоционального стресса может проявляться в двух формах: возбудимой,
которая характеризуется повышенной двигательной активностью, суетливостью,
излишней говорливостью и т. д., и тормозной, для которой характерны
угнетение деятельности, замедление реакции, неспособность отвечать на различные
воздействия.
Положительное влияние
эмоционального стресса проявляется в мобилизации мозговой активности. Особенно
отчетливо это выражено в условиях, когда необходимо прогнозировать вероятный
исход своих действий и, соответственно, вероятность появления тех или иных
сигналов.
Исследования
прогностической деятельности человека в вероятностно организованной среде
показывают, что существует несколько принципиально различных тактик работы, а
успешность деятельности в значительной мере зависит от уровня мотивации и
психофизической напряженности оператора.
Основные требования к
условиям труда человека—оператора в сжатой форме сводятся к трем следующим
условиям. 1. Тщательный профессиональный отбору и хорошая подготовка оператора
к данному виду деятельности. 2. Детальный учет человеческого фактора при
проектировании оборудования и технических систем. 3. Учет экологического
фактора, связанного с социальной средой обитания человека и с непосредственными
условиями труда.
3.25. ЦЕНТРАЛЬНАЯ РЕГУЛЯЦИЯ ДВИЖЕНИЙ
Все многообразие видимых форм
движений животных и человека основывается на физических законах перемещения тел
в инерциальном пространстве. При классификации движений необходимо
учитывать конкретные целевые функции, которые должна выполнять двигательная
система. В самой общей форме таких функций четыре: 1) поддержание определенной
позы; 2) ориентация на источник внешнего сигнала для его наилучшего восприятия,
а также поддержания равновесия тела; 3) перемещение тела в пространстве; 4)
манипулирование. Иерархия уровней мозгового управления движениями также
находится в зависимости от требований к структуре движения. Если подкорковый
уровень связан с набором врожденных или автоматизированных программ, то корковый
уровень организует спинномозговой моторный аппарат для выполнения произвольных
и тонких движений.
3.25.1. Управление ориентационными движениями и позой
Ориентационные движения. Их система связана с ориентацией
тела относительно координат внешнего пространства и с установкой сенсорных
аппаратов в положение наилучшего восприятия внешнего стимула. Примером первого
может служить функция поддержания равновесия, второго — движения фиксации
взора.
При ориентации в пространстве относительно зрительных координат
одной из основных задач является фиксация взора, которая выполняется в основном
глазодвигательной системой. Изображение неподвижного или движущегося предмета
фиксируется в наиболее чувствительном поле сетчатки. Точной фиксации глаз или
стабильному их перемещению по определенной траектории противодействуют
различные помехи: движения головы, самого зрительного объекта и изменения
глазодвигательной системы. Компенсация этих помех достигается с помощью
специальных рефлекторных механизмов.
Быстрые скачки глаз (микросаккады), которые служат
для переноса точки фиксации, делят на произвольные и непроизвольные. Произвольные
саккады отражают сознательно запрограммированную смену фиксаций взора в
зависимости от общего плана исследовательского поведения и не зависят от характеристик
объекта. Непроизвольные саккады участвуют в механизмах фиксации,
корректируя отклонения сетчатки от нужного положения, возникающие вследствие
дрейфа глаз или смещения головы. Они регулируются характеристиками зрительного
объекта и сигналами от глазных мышц. Непроизвольные саккады могут служить для
квантования зрительного мира, стирая остаточные следы предыдущего раздражения
сетчатки. Каждый видимый объект точно локализован в пространстве по отношению к
некоторому «образу тела». Простое движение (например, быстрое касание
какого—либо предмета) полагает, что при формировании команды учитываются
положение цели на сетчатке, положение глаз относительно координатной системы и
положение головы относительно тела.
В организации непроизвольных саккад большую роль выполняет мозжечок,
который участвует в программировании таких быстрых движений, которые не
могут регулироваться по кольцу внешней обратной связи, т. е. с помощью
проприоцептивной афферентации. Однако это не исключает возможности
регулирования по «внутренней» обратной связи.
Ведущую роль в управлении произвольными саккадами играет кора
больших полушарий. Для затылочного глазодвигательного поля определяющими
будут
|
|
|
Рис. 3.63 Соматотопическая
организация корковой зоны SI человека Изображения
под поперечным срезом мозга (на уровне постцентральной извилины) и их
обозначения демонстрируют пространственное представительство поверхности тела
в коре, установленное путем локальной электрической стимуляции мозга
бодрствующих больных. |
параметры зрительного сигнала, а для
фронтального и теменного глазодвигательных полей — сигналы от мозжечка,
связанные с выполнением «внутренней» двигательной программы. Предполагают, что
глазодвигательная система вовлекается в работу не после полного описания
изображения в зрительной сенсорной системе, а непосредственно по ходу обработки
зрительной информации.
Координированное движение глаз и головы регулируется
специальной системой рефлексов: Например, при слежении за целью повороты
головы целиком определяются движением глаз, а при поиске источника звука в
темноте одновременно запускается движение глаз и головы к цели. При появлении
на свету неожиданного зрительного объекта вначале запускается саккадическое
движение к цели и спустя некоторое время начинается поворот головы в том же
направлении. Центральным звеном координированного движения глаз и головы
являются верхние холмики пластинки крыши среднего мозга и система шейных,
глазных и вестибулярных рефлексов.
Управление позой. Позу тела можно описать как совокупность некоторых
значений суставных углов, обладающих определенной ориентацией в поле тяготения.
Механизмы позы распадаются на две категории: фиксацию определенных положений
тела и конечностей, т. е. фиксацию движения в суставах, и ориентацию сегментов
тела относительно внешних координат, т. е. поддержание равновесия. Исходная
поза тела накладывает некоторые ограничения на последующее движение.
К низшим механизмам управления позой
относятся спинальные рефлексы, шейные установочные рефлексы и вестибулоспинальные
рефлексы. К высшим механизмам управления позой относятся механизмы формирования
«схемы тела».
Термином схема тела обозначают систему обобщенной
чувствительности собственного тела в покое и при движении, пространственных
координат и взаимоотношений отдельных частей тела.
Детальное представительство отдельных частей тела, вплоть до
фаланг пальцев (соматотопия), характерно для соматосенсорной системы. Общая
«карта» тела для каждого полушария мозга представлена в виде «гомункулюса»
(рис. 3.63). Чувствительность всего тела, топографически распределенная по
поверхности коры, составляет ту основу, тот исходный материал, из которого
путем объединения формируются целостные функциональные блоки крупных отделов
тела. Эти интегративные процессы завершаются у взрослого организма и
представляют собой закодированное описание взаиморасположения частей тела,
которые используются при выполнении автоматизированных стереотипных движений.
|
|
|
Рис. 3.64 Высшие
механизмы регуляции позы. (Объяснение
см. в тексте). |
Базой для этих процессов служит анатомически закрепленная
«карта» тела, поэтому они составляют лишь основу статического образа тела.
Для его формирования необходимо соотнести эту информацию с положением тела по
отношению к силе земного притяжения и взаиморасположением функциональных блоков
тела в системе трех пространственных плоскостей. Вестибулярная система
воспринимает перемещения всего тела вперед—назад, вправо—влево, вверх—вниз, а
соответствующая информация поступает в теменные области коры мозга, где
происходит ее объединение с информацией от скелетно—мышечного аппарата и кожи
(рис. 3.64). Туда же поступает импульсация от внутренних органов, которая также
участвует в создании на бессознательном уровне статического образа тела.
Таким образом, статический образ тела представляет собой
систему внутримозговых связей, основанную на врожденных механизмах и
усовершенствованную и уточненную в индивидуальной жизни.
Осуществляя тот или иной род деятельности, человек меняет
взаиморасположение частей тела, приобретает в порядке научения новые
двигательные навыки, а значит, формирует новые трехмерные пространственные
модели тела, т. е. динамический образ тела.
В отличие от статического он имеет значение лишь для данного
конкретного момента времени, для определенной ситуации, при изменении которой
он сменяется новым. Динамический образ базируется на информации, поступающей от
кожи, мышц, суставов и вестибулярного аппарата.
В мозгу происходит постоянное взаимодействие того и другого
образов тела, осуществляется сличение динамического образа с его статическим
аналогом. В результате этого формируется субъективное ощущение позы,
отражающее не только положение тела в данный момент, но и возможные его
изменения в непосредственном будущем. Если согласование не достигнуто, то
вступают в действие активные механизмы перестройки позы. Итак, для того чтобы
субъективно оценить позу, необходимо сопоставить закодированный в памяти эталон
статического образа тела с его любой конкретной вариацией — динамическим
образом тела.
Не только схема тела является трехмерно пространственной, и
сам образ реальной пространственной ситуации воспринимается в том же
трехкоординатном пространственном измерении. Человек оценивает окружающее
пространство не абстрактно, а применительно к самому себе, т. е. включает свое
тело в качестве непременного компонента пространства.
Человек «вписывается» в окружающую ситуацию и воспринимает
ее, «привязывая», соотнося внешние объекты с положением собственного тела.
Внутренний и внешний образы описываются одним и тем же физиологическим языком,
в котором закодирован весьма ограниченный набор одних и тех же параметров.
Для подробного описания нужно представить себе наличие,
например, какой—то вертикальной плоскости, которая делит тело на две
симметричные половины. Тогда все участки тела и предметы окружающей среды,
расположенные
|
|
|
Рис. 3.65 Формирование
представлений о внешнем пространстве и «схеме тела» Объяснение см. в
тексте. |
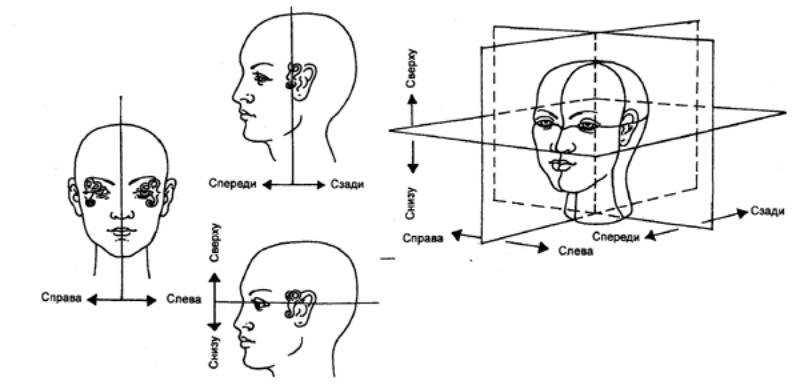
по одну сторону от этой плоскости, будут
субъективно восприниматься как «слева», а по другую — как «справа» (рис. 3.65).
Только информация о местоположении органов тела будет передаваться по
соматическим трактам, а информация о местоположении предмета по отношению к
организму — по зрительным трактам.
Эти два потока информации
встречаются и объединяются для последующего использования в произвольной
двигательной деятельности. Конвергенция и интеграция этих афферентных влияний
происходят в пределах таламопариетальной ассоциативной системы мозга.
Рассмотренное справедливо не только для описанной выше сагиттальной плоскости,
но и для фронтальной, которая проходит через центры вестибулярных аппаратов, и
горизонтальной, которая проходит через центры глазных яблок и вестибулярных
аппаратов.
Таким образом, и наклон головы, и
содружественный поворот глазных яблок в ту или иную сторону от этих плоскостей
способствуют восприятию внешнего и внутреннего пространства в одних и тех же
параметрах. В итоге создается целостный образ организма в среде, который служит
определенным сенсорным фоном для выполнения любого произвольного движения.
Под термином локомоция (от
лат. locus — место и motio — движение) понимают перемещение тела из одного места
пространства в другое, для чего необходимы некоторые силы, изменяющие исходное
состояние организма. Развиваемые при этом усилия должны преодолеть силу
тяжести, сопротивление окружающей среды и силы инерции тела. На локомоцию
влияют характер и рельеф местности. Во время локомоции необходимо непрерывно
поддерживать равновесие.
Наиболее распространенным примером
локомоции являются ходьба и бег, которые отличаются стереотипным движением
конечностей, причем для каждой из форм локомоции характерны две фазы шага:
опоры и переноса. Разные животные обнаруживают различные, свойственные только
им способы ходьбы, которые в то же время обладают общими чертами. Локомоция
характеризуется
|
|
|
Рис. 3.66 Модель спинального генератора
локомоции Приведены афферентные и эфферентные связи на спинальный генератор;
Ф — флексоры (сгибатели), Э —
экстензоры (разгибатели), α и γ — альфа— и гамма—мотонейроны. |
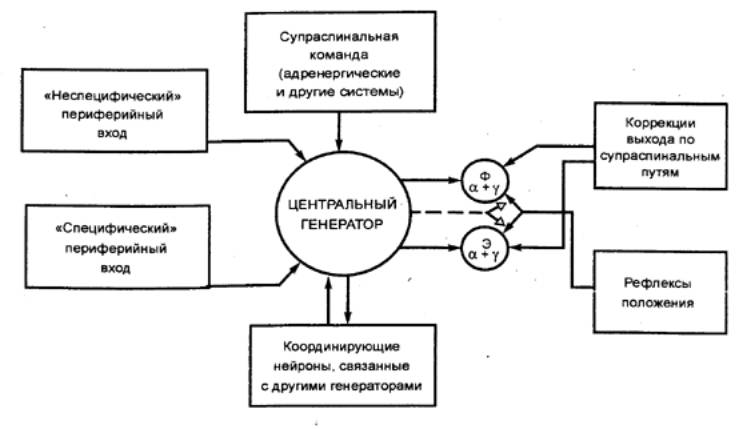
походкой — определенным способом передвижения
по поверхности, который оценивается по распределению во времени циклических
движений конечностей, длительностью опорной фазы и последовательностью
перемещения опорных конечностей.
Описана нейронная цепь, находящаяся в спинном мозгу, которая
называется спинальным генератором шагания. Она ответственна за
чередование периодов возбуждения и торможения различных мотонейронов и может
работать/в автоматическом режиме (рис. 3.66). Элементарной единицей такого
центрального генератора является генератор для одной конечности. Допускают
даже, что мышцы, управляющие одним суставом, могут контролироваться каким—то
собственным унитарным генератором. При формировании локомоторных движений эти
генераторы разных движений тесно связаны и оказывают друг на друга возбуждающее
влияние.
Структуры спинного мозга находятся под непрерывным супраспинальным
контролем. Установлено, что раздражение определенных участков ствола мозга
способно вызывать координированную локомоцию, обусловленную запуском
ритмической активности спинального генератора.
Супраспинальный контроль локомоции выполняет несколько
основных функций: 1) быстро запускает локомоцию, поддерживает постоянную
скорость или изменяет ее, если требуется, а также прекращает ее в нужный
момент; 2) точно адаптирует движение (и даже отдельный шаг) к условиям среды;
3) обеспечивает достаточно гибкую локомоторную позу, чтобы соответствовать
различным условиям передвижения, таким, например, как ползанье, плаванье, бег
по снегу, перенос груза и т. д. Афферентная сигнализация, идущая в спинной мозг
от мышечных веретен, сухожильных рецепторов Гольджи, кожи не является необходимым
условием возникновения локомоции, тем не менее она участвует в более точной
адаптации локомоторных движений к условиям среды.
Значительную роль выполняет мозжечок, который
обеспечивает коррекцию и точность постановки конечностей на основе сравнения информации
о работе спинального генератора и реальных параметров движений, причем
мозжечковые влияния направлены на регуляцию моторного выхода, а не на
активность самого спинального генератора. Мозжечок получает прямую информацию
по восходящим спинно—мозжечковым путям. Предполагают, что мозжечок
программирует каждый следующий шаг на основе информации о предыдущем.
|
|
|
Рис.
3.67 Соотношение
прямых и внутренних обратных связей в ядрах дорсальных столбов 1 — мышца, 2 — чувствительный нейрон спинального ганглия, 3 — мотонейрон
спинного мозга, 4 — спинной
мозг, 5 — интернейрон спинного мозга, 6
— нейрон красноядерно—спииномозгового пути, 7 — красное ядро, 8 —
пирамидный путь, 9 — ядра
дорсальных столбов, 10 —
нейроны продолговатого мозга, дающие начало медиальной петле (11), которая переключается в
таламических ядрах и несет афферентацию в кору мозга. На этих нейронах (10) осуществляется
пресинаптическое торможение в момент эфферентного залпа по коллатералям
волокон пирамидного пути; последние берут начало от крупных пирамидных клеток
(12} сенсо—моторной области
коры (13). |
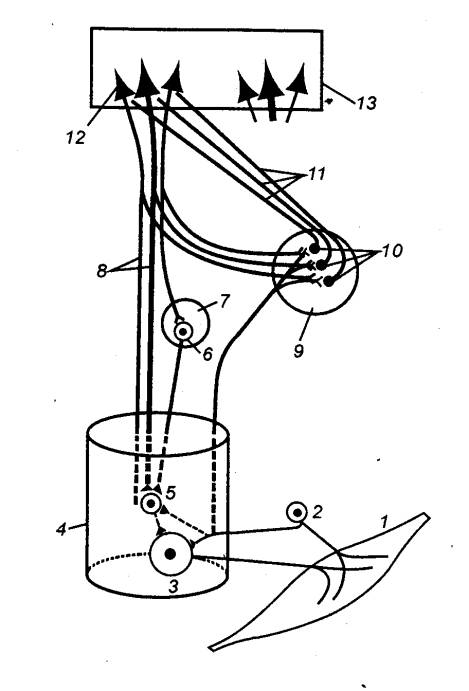
Другой важнейший уровень мозга, куда направляется информация
о характере выполнения движения, это полушария мозга с таламическими ядрами,
стриопаллидарной системой и соответствующими зонами коры головного мозга. В
быстрых движениях баллистического типа классическая соматическая обратная связь
не принимает участия потому, что само движение завершается раньше, чем успеет
поступить информация от двигательных аппаратов к соответствующим мозговым
центрам. Медленные движения постоянно корректируются благодаря показаниям с
соответствующих сенсорных датчиков, расположенных в скелетно—мышечном аппарате
и передающих информацию в разные отделы мозга вплоть до коры. Эти сенсорные
коррекции способны изменить характер движения по ходу его осуществления.
Движения, базирующиеся на врожденных координациях, в меньшей степени требуют
обратной связи от моторного аппарата, а движения, в основе которых лежит
формирование новых координационных отношений, всецело зависят от соматической
афферентации с двигательного аппарата. Кроме того, степень участия обратных
связей находится в зависимости и от уровня автоматизации двигательного акта.
Простые движения, осуществляющиеся без
проприоцептивной обратной связи (саккады и быстрые движения конечностей),
выполняются по жесткой «запаянной» программе. Любое же сложное движение требует
предварительного программирования. Для сложных движений существенным является
сличение обратной афферентации с тем сенсорным образом движения, который
формируется в составе программы. Результат сличения и является основным
фактором перестройки программы. Эти влияния передаются к аппаратам
программирования по каналам внутренней обратной связи, которая включает в себя
все процессы перестроек двигательной программы в зависимости от
внутрицентральных влияний.
Такими каналами внутренней обратной связи являются медиальные
петли (лемниски), по которым передается импульсация, интегрированная в
ядрах дорсальных столбов. Местом ее поступления в кору больших полушарий
оказываются не только соматосенсорные зоны, но и сенсомоторные области, куда,
кроме того, конвергирует гетеромодальная информация и где осуществляются
интегративные процессы.
Если перемещение конечности — это компонент
общекоординированного акта, то внутренняя (проприоцептивная) и внешняя (кожная
и телецептивная) афферентации выступают как содружественные звенья системы
сенсорного обеспечения движений. Соматическая афферентация от локомоторного
аппарата необходима для работы генератора инициации движений и их
координированного выполнения. Это в первую очередь касается медленных
компонентов двигательных актов. Существует обратная связь, с помощью которой
кора информируется не об отдельных параметрах движений, а о степени
соответствия предварительно созданной двигательной программы тому наличному
движению, которое достигается в каждый момент времени. Одним из важнейших
каналов такой внутренней обратной связи и выступают медиальные петли.
Известно, что пирамидный путь отдает
большое число коллатералей к ядрам дорсальных столбов в продолговатом мозгу.
Поэтому «пирамидальный залп» из коры достигает не только аппаратов спинного
мозга, но и ядер дорсальных столбов, нейроны которых в течение реализации
данного цикла движения находятся в тормозном состоянии благодаря развитию в них
пресинаптического торможения (рис. 3.67).
Следовательно, импульсация по
волокнам дорсальных столбов (тонкий и клиновидный пучки) оказывается
заблокированной, т. е. подпороговой. Превращение в надпороговую при отклонении
параметров реального движения от требуемых согласно команде вызывает поток
импульсов по системе медиальных петель в кору. Здесь будет проводиться
коррекция следующего цикла командного воздействия. Поэтому сенсомоторная кора →
коллатерали пирамидного пути → ядра дорсальных столбов →медиальные
петли — это важнейшее кольцо, по которому происходит непрерывная коррекция
двигательных команд сообразно параметрам предшествующего двигательного цикла.
Срочная соматическая информация
обеспечивает фильтрацию телецептивных и экстралемнисковых влияний на
корково—подкорковом уровне путем управления системой нисходящих связей. Этим
достигается организация точных выходных команд, придающих движению
координированный характер. Ядра дорсальных столбов являются одной из тех
мозговых структур, где осуществляется сличение результата двигательного цикла с
копией программы и последствия рассогласования поступают в сенсомоторную кору
для очередной коррекции двигательной программы. Следовательно, если внешняя
обратная связь от соматических рецепторов ограничивается спиннобульбарным
уровнем, то внутренняя обратная связь осуществляется по системе медиальных
петель.
3.25.3. Организация манипуляторных
движений
Манипуляторные движения — это наиболее яркий пример
произвольных движений, возникновение которых обусловливают внутренние
мотивационные влияния. Эти движения являются локальными, что ставит перед
двигательной системой следующие задачи: 1) выбор ведущего мышечного звена; 2)
компенсацию внешней нагрузки; 3) настройку позы; 4) соотнесение координат цели
и положения собственного тела.
Отличительной чертой манипуляторных движений является их
зависимость от центральной программы и телецептивных сенсорных входов. Поэтому ведущая
роль отводится следующим мозговым структурам: лобной коре, базальным ядрам
и мозжечку. Активность других супраспинальных цепей двигательного контроля в
большей степени зависит от периферической информации, поступающей вслед за
реализацией команд.
Ведущая роль в программировании быстрых движений принадлежит
мозжечковой системе, а в программировании медленных — базальным
ядрам. Выходы этих структур на спинальный уровень опосредованы в основном
через сенсомоторную кору, которая дополнительно к этому обладает своими собственными
функциями.
3.25.4. Корковая сенсомоторная
интеграция
Структурной единицей неокортекса
является модуль — микросистема тесно контактирующих между собой
пирамидных нейронов для их синхронного вовлечения в формирование эфферентного импульсного
залпа (рис. 3.68). Такое объединение рассматривают в качестве жесткого
структурного элемента организации коры, формирующегося в пренатальном
онтогенезе. Почти всем модулям свойственна широкая конвергенция гетеросенсорных
влияний. В моторную кору конвергируют те компоненты сенсорной информации,
которые являются жизненно важными для организма. Это информация о биологической
значимости того или иного сигнала либо его значимости, приобретенной в порядке
выработки условного рефлекса.
Топографическое распределение
соматических проекций участков тела на кору больших полушарий только частично
определяется анатомическими закономерностями. Не менее важную роль играют
функциональные механизмы, определяющим среди которых является внутрикорковое
торможение.
Структурный корковый
модуль выступает в качестве элементарной сенсо—моторной единицы. Различия между
отдельными модулями определяются характером соматической сенсорной проекции и
адресатом эфферентного выхода из того же модуля. Существует распределенная
система корковых модулей, выходы которых конвергируют на разных уровнях
ствола и спинного мозга, обеспечивая управление деятельностью определенной
мышечной группы. Распределенная система является частью более обширных
общемозговых систем, сформированных в раннем онтогенезе. Такие функциональные
структуры управляют более обширной частью скелетно—мышечного аппарата.
|
|
|
Рис. 3.68 Функциональные объединения нейронных
модулей Одиночный
структурный модуль имеет вид цилиндра; 4—6 модулей могут сгруппироваться в функциональные
концентрические объединения, между которыми . возможны как облегчающие, так и
взаимотормозные отношения. Обширные функциональные объединения из нескольких
концентрических модулей строятся на основе ветвлений аксонной коллатерали
пирамидных нейронов (1),
ветвлений таламического афферента (2)
и ветвлений аксона корзинчатой клетки (3); стрелками показано
направление восходящей и нисходящей импульсации. |
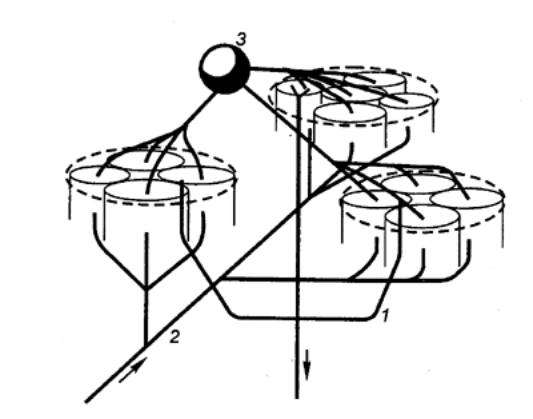
Вовлечение корковых систем в целостную деятельность мозга
определяется текущей мотивацией и конкретной двигательной программой,
направленной на удовлетворение этой мотивации. Двигательная программа, чтобы
носить приспособительный характер, должна учитывать все сигнально значимые
компоненты внешней среды, относительно которых совершается целенаправленное
движение, т. е. строиться на принципе мультисенсорной конвергенции.
Телецептивные входы на модулях сенсомоторной коры носят сравнительно диффузный
характер и адресуются ко многим пространственно разнесенным одноименным
модулям. Констелляции модулей основаны на синергичных и антагонистичных
межмодульных соотношениях, что обеспечивает динамичность их участия в различных
актах сенсомоторной интеграции.
3.25.5. Программирование движений
Основным элементом любого
целенаправленного движения является формирование предварительных программ, т.
е. процессов, позволяющих предвидеть динамику изменений факторов внешней среды
и придать будущему движению адаптивный характер. Результат сличения
афферентации о движении с двигательной программой, передающийся по системе
внутренней обратной связи, является основным фактором перестройки программы.
Последнее зависит от биологической мотивированности движения, его временных
параметров, моторной дифференцированности, сложности координационного состава и
уровня его автоматизированное.
Мотивации определяют общую стратегию
движения. Каждый конкретный двигательный акт рассматривается как шаг к
удовлетворению мотивации, т. е. решает некоторую промежуточную задачу или цель
(рис. 3.69). Биологические мотивации приводят к запуску либо «запаянных»,
жестких моторных программ, либо формируют новые сложные программы. Если
мотивация определяет цель движения, его программу, то она же обусловливает ее
зависимость от сенсорных факторов. В качестве обратной связи здесь выступает
удовлетворение мотивации. Тактика движения, или двигательная команда,
определяет, как будет осуществляться требуемое запрограммированное движение, т.
е. каково распределение во времени тех эфферентных залпов к моторным ядрам
спинного мозга, которые определяют активацию различных мышечных групп.
В отличие от программы команды
движения должны быть адаптивными к функциональному состоянию самого
скелетно—двигательного аппарата как непосредственного исполнителя этих команд.
Предполагают, что срочная идентификация состояния мышечной системы
осуществляется путем оценки реакции
|
|
|
Рис.
3.66 Модель спинального генератора локомоции Приведены афферентные и
эфферентные связи на спинальный генератор; Ф — флексоры (сгибатели), Э — экстензоры (разгибатели), α
и γ — альфа— и гамма—мотонейроны. |
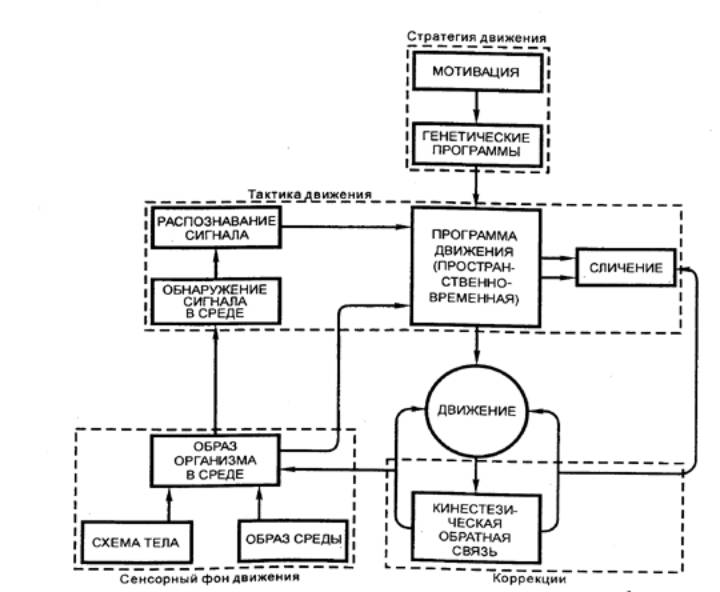
системы на небольшое точно
дозированное возмущение (тест—импульс), которое подается непосредственно
перед или с момента запуска команд.
Тест—импульс подается только на те мышцы, которые должны будут
участвовать в следующем движении. Реакция мышц будет не настолько значительна,
чтобы существенно изменить само движение, но достаточна для возбуждения
мышечных веретен при изменении длины интрафузальных волокон на 5—10 мкм.
Информация о реакции на тест—импульс конвергирует на некотором промежуточном
уровне с основным управляющим воздействием (командой), которое корректируется в
соответствии с состоянием самого двигательного аппарата. Такое тестовое
воздействие передается по быстропроводящим волокнам пирамидного пути и
адресуется к сегментам спинного мозга.
Непосредственное управление движением обусловливается
активностью сенсомоторной коры мозжечка и полосатого тела. Последнее участвует
в трансформации «намерения действовать» в соответствующие «командные сигналы»
для инициации и контроля движений.
Ассоциативные системы мозга и движение. Специфичность участия таламопариетальной
ассоциативной системы в организации движений определяется двумя моментами:
она участвует в формировании интегральной схемы тела, все части которого
соотнесены не только друг с другом, но и с вестибулярными и зрительными
сигналами, и регуляции внимания к текущим сигналам окружающей среды с учетом
ориентации всего тела относительно этих сигналов. Эта система «привязана» к
настоящему моменту времени и к анализу пространственных взаимоотношений
разномодальных признаков.
По таламофронтальной ассоциативной системе передается информация о
мотивационном состоянии и сопутствующих висцеральных изменениях. Фронтальная
ассоциативная кора опосредует мотивационные влияния на организацию поведения в
целом благодаря связям с теменно—височной корой, гипоталамусом, областями
среднего и промежуточного мозга. Поэтому лобная кора участвует в регуляции
процессов во внутренней среде организма, сенсорных и моторных механизмах мозга.
3.25.6. Функциональная структура произвольного движения
Среди общих принципов управления
движениями выделяют проблему организации единовременной команды к
исполнительным аппаратам. Если ее рассматривать безотносительно к стратегии и
тактике движения, то основной целью системы, формирующей эту команду, будет
координация всех компонентов команды.
Для того чтобы выполнить эту задачу, система должна
осуществлять следующие процессы: идентификацию состояния системы, определение
целевой функции ведущего кинематического звена, подстройку соотношения
активности мышц—антагонистов. Последнее производится автоматически благодаря
системе спинальных рефлексов. Идентификация же состояния двигательной системы
обеспечивается двумя путями: оценкой реакции системы на тест—импульс,
генерируемый корой, или на внешнее воздействие.
ЦНС располагает значительным числом генетически закрепленных
программ типа локомоторной программы, базирующейся на активности спинального
генератора шагания. Такие простые программы объединяются в более сложные
системы типа поддержания вертикальной позы, что происходит в процессе научения
при участии структур переднего мозга.
Одной из задач управления движением является выбор ведущего
кинематического звена. Этот выбор предопределен исходной позой, перестройка
которой всегда опережает локальное движение. Информация об исходной позе будет
перераспределять активность сенсомоторной коры. Подкорковые управляющие
структуры (полосатое тело и мозжечок) формируют новое движение почти исключительно
путем модификации состояния командных интернейронов во врожденных подсистемах
управления. Диапазон этой модификации очень широк и у многих видов животных
объемлет почти весь двигательный репертуар.
Для формирования новых координационных отношений, особенно в
случае манипуляторных движений, необходимо подавить активность командных
нейронов врожденных программ и сформировать параллельный управляющий канал.
Таковым является корково—спинномозговой путь, степень развития которого
независимо от положения животного в филогенетическом ряду определяет роль
манипуляторных движений в общем двигательном поведении. Морфологической основой
организации тонких манипуляторных движений является сенсомоторная кора с ее
структурными модулями и объединениями модулей разного порядка сложности. Все
элементы вышепоименованных систем одномоментно связаны между собой, поэтому
изменение любого звена влечет за собой сдвиг состояния всей двигательной
системы в целом.
Ориентационные движения и следящие движения глаз, головы,
конечностей представляют собой задачу более высокого порядка, чем координация
движений сегментов тела. Эти движения зависят от функционирования таламопариетальной
ассоциативной системы мозга.
Самой сложной, филогенетически самой молодой является
способность формировать последовательность движений и предвидеть ее реализацию.
Решение этой задачи связано с фронтальной ассоциативной системой,
которая запоминает и хранит в памяти такие последовательности движений.
Высшим отражением этого кодирования у человека является
вербализация основных понятий движения.
Одно из основных проявлений поражений лобных долей человека
состоит в нарушении грамматической структуры речи и невозможности использования
вербальных команд построения последовательности движений.
Всеобщей закономерностью работы системы управления
движениями является использование обратной связи. Сюда входит не только
проприоцептивная обратная связь от начавшегося движения, но и активация систем
поощрения или наказания. Кроме того, включается и внутренняя обратная связь, т.
е. информация об активности нижележащих уровней двигательной системы или
эфферентная копия самой двигательной команды. Этот вид обратной связи необходим
для выработки новых двигательных координации. Для движений разной сложности и
скорости обратная связь может замыкаться на разных уровнях. Поэтому оба типа
управления — программирование и слежение — могут сосуществовать в системе
управления одним и тем же движением.
3.26. ЭМОЦИИ КАК КОМПОНЕНТ ЦЕЛОСТНЫХ ПОВЕДЕНЧЕСКИХ РЕАКЦИЙ
Различные формы поведения человека и
животных направлены на удовлетворение многообразных потребностей, лежащих в
основе поддержания индивидуального и видового гомеостаза. Объективное описание
целостных поведенческих актов и сопутствующих им вегетативных реакций,
исследование центральной организации взаимоотношений организма с внешним миром,
возможность прогнозировать ответы на изменения во внутренней среде и на сигналы
из внешнего мира — необходимая основа физиологического осмысления механизмов
высшей нервной деятельности. Нельзя упускать из виду и процессы, отражающие
субъективные состояния, но поддающиеся объективному нейрофизиологическому
анализу.
К таким состояниям следует отнести
прежде всего мотивации и эмоции. Хотя представления о них пришли
в физиологию из психологии, современные исследователи не могут обойтись без их
использования при анализе механизмов высшей нервной деятельности. О мотивационно—эмоциональной
сфере человека судят по его поступкам, мимике и другим выразительным
движениям, вегетативным реакциям, электрическим явлениям в головном мозгу, а
также по словесному отчету о субъективном состоянии и собственному отношению к
нему.
3.26.1. Биологическая роль эмоций
Судить о мотивационно—эмоциональных
процессах у животных можно лишь по объективным показателям. Однако накопленные
сведения об эволюции головного мозга, сопоставление участия отдельных его
систем в организации поведения человека и животных дают возможность
постулировать существование у последних субъективных состояний как сформировавшихся
в процессе эволюции полезных признаков, необходимых для выживания особи и
сохранения вида.
В целостной поведенческой реакции
мотивация и эмоция проявляются в неразрывном единстве, но, как показывает
физиологический анализ, в экспериментальных условиях их можно разделить, так
как они отражают активность хотя и тесно взаимодействующих, но
специализированных отделов ЦНС. Особенно четко демонстрируется относительная
самостоятельность мотиваций и эмоций при разрушении этих отделов или
раздражении их электрическим током или фармакологическими веществами.
Мотивации и эмоции тесно связаны с
возникновением и удовлетворением потребностей организма — необходимым условием
его жизнедеятельности.
Мотивации (побуждения, влечения, драйвы)
определяются генетической программой и индивидуальным опытом, активизируются
потребностями и проявляются в избирательном возбуждении специализированных
мозговых структур, что приводит при благоприятной внешней ситуации к реализации
целенаправленного поведения.
Различают низшие (первичные,
простые, биологические) и высшие (вторичные, сложные, социальные)
мотивации. Примерами биологических мотиваций могут служить голод, жажда, страх,
агрессия, половое влечение, забота о потомстве. Не следует отождествлять
мотивации и потребности. Нередко последние не трансформируются в мотивационное
возбуждение и, следовательно, не могут быть удовлетворены посредством
реализации целенаправленного поведенческого акта.
Крысы, например, предпочитают жирную
безбелковую пищу безжировой белковой. Люди нередко также потребляют менее
полезную, а иногда и вредную пищу с высоким содержанием низкомолекулярных
Сахаров лишь потому, что она вкуснее, чем оптимальная по составу. У человека и
многих видов животных нет врожденного предпочтения питательных веществ, богатых
витаминами. Поэтому на протяжении многих веков отдельные группы населения
страдали болезнями, вызываемыми авитаминозами (цинга, пеллагра, бери—бери и
др.), причем развитие этих болезней не всегда определялось отсутствием
источника витаминов.
Вместе с тем для удовлетворения
многих биологических потребностей не обязательно существование врожденных
механизмов их трансформации в мотивационное возбуждение. Специализированные
поведенческие акты могут формироваться на основе неспецифического поиска,
инициируемого различными нарушениями гомеостаза. В частности, у крыс могут быть
выработаны витаминные, минеральные и другие аппетиты, а также предпочтение или
отвергание определенных пищевых продуктов. Подобные перестройки носят
условнорефлекторный характер и свидетельствуют о большой пластичности
биологических мотиваций. Естественно, в еще большей мере связаны с
индивидуальным опытом высшие, социальные мотивации.
Перестройки поведения
требуют оценки произведенных действий. Она основана на формировании
положительного или отрицательного эмоционального состояния. Первое отражает
благотворный эффект произведенного действия и создает предпосылку для его
продления или повторения, второе сигнализирует о вредном его влиянии на
организм и нецелесообразности повторения. Эмоции тесно связаны с подкреплением
— важнейшим компонентом обучения.
Таким образом, в
поведении отражаются отдельные, но тесно переплетающиеся явления. На основании
потребностей формируется мотивация, ее реализация в целенаправленных актах
поведения сопровождается эмоциями положительного или отрицательного знака.
Среди этих явлений
наиболее сложны для изучения именно эмоции. Анализ потребностей можно проводить
на основании консуматорных реакций. Так, потребность в воде или пище
определяется по их потреблению. Уровень биологической мотивации измеряется
многими способами, в частности скоростью целенаправленной побежки, преодолением
препятствий разной сложности, выбором действий в условиях конкурентных
отношений между удовлетворением разных биологических потребностей (пищевой и
половой, пищевой и водной и т. п.). В естественных условиях мотивации
проявляются как эмоционально окрашенные состояния, однако параллелизма между
уровнями мотивационного и эмоционального возбуждения не имеется. Поэтому нельзя
оценивать последнее по актам, нацеленным на удовлетворение биологических
потребностей.
Важной особенностью
эмоций является комплекс телесных реакций, сопутствующих отрицательным
или положительным переживаниям. У человека субъективные состояния удовольствия
или неудовольствия находят внешнее выражение в изменениях позы, походки,
выражения лица, жестов, голосовых реакций, частоты пульса, дыхания,
потоотделения, многих гуморальных показателей. По этим изменениям можно
косвенно судить о степени волнения испытываемого человеком. Однако условность
такого суждения очевидна. Мимические и прочие движения, характерные для
аффективных состояний, человек в состоянии затормозить или воспроизвести при
отсутствии соответствующих переживаний.
Во многих случаях внешние выражения чувств определяются социальными
стереотипами поведения. Тренировка позволяет подавлять или активизировать
некоторые вегетативные реакции, отражающие эмоциональные переживания. Наиболее
полно такая способность проявляется в игре актеров.
Таким образом, даже у человека на основании сопоставления
словесного отчета о субъективном состоянии и комплекса моторно—вегетативных
реакций можно лишь догадываться об испытываемых им переживаниях.
Еще более сложно оценить эмоциональное состояние у животных.
О нем судят по различным реакциям на действие внешних раздражителей или
стимуляцию некоторых отделов головного мозга. Вместе с тем общепринятое
положение о единых для человека
и животных закономерностях эволюции мозга позволяет считать, что первичные
эмоции сформировались задолго до появления человека как чувственное выражение
инстинктивной деятельности.
Именно с таких позиций
рассмотрел их Ч. Дарвин в работе «О выражении эмоций у человека и животных»
(1872). Им был приведен большой материал по сравнительному анализу мимики,
пантомимики, голосовых и вегетативных реакций при аффективных состояниях у
представителей разных отрядов млекопитающих и человека. Выразительные движения
рассматривались Ч. Дарвином как сформировавшиеся в процессе естественного
отбора приспособительные реакции, существенные для общения с особями своего или
других биологических видов. Он предположил, что мимико—жестикуляционные реакции
являются компонентами агрессивного, оборонительного или других форм поведения.
Например, отражающие аффект гнева оскаливание зубов и угрожающие жесты пугают
врага. Некоторые выражения аффективного состояния являются рудиментами. Так, в
состоянии гнева человек сжимает кулаки и скалит зубы, показывая готовность к
борьбе.
Ч. Дарвин оценивал
эмоциональные состояния в связи с побуждением к действию, не пытаясь
анализировать их субъективную сторону. Подобный подход был полезен для
подтверждения сходства в поведении животных и человека и обоснования
происхождения человека от обезьяны. Однако при этом игнорировались качественные
различия между психической сферой человека и животных.
3.26.2. Эмоции и психическая
деятельность
Периоду естественнонаучного подхода
к анализу эмоций предшествовало длительное осмысление их места в психической деятельности
человека. Еще в XVII в. Р. Декарт выдвинул положение, согласно которому под
влиянием эмоции организм осуществляет полезные для него действия и уклоняется
от совершения вредных. В работах философов XVIII — начала XIX вв. большое место
занимал вопрос о взаимоотношении аффективной сферы и телесных реакций.
Высказывались предположения о зависимости эмоциональных состояний от
органических изменений и о противоположных причинно—следственных отношениях.
Важно отметить, что некоторые философы пришли к заключению о связи эмоций с
конфликтом между представлениями (т. е. в соответствии с современной нам
терминологией — рассогласованием).
В конце XIX в. американский психолог
и философ У. Джеме предпринял попытку создания физиологической теории эмоций,
вторичных, по его мнению, по отношению к вызванным различными стимулами
телесным изменениям. Согласно «периферической теории» У. Джемса, эмоция
является осознанием изменений в работе внутренних органов и мышц. Близкую
позицию занял датчанин К. Ланге, полагавший, что эмоциональное состояние
отражает органические изменения, связанные преимущественно с кровообращением.
Вопреки представлению об эмоции (психическом состоянии) как первопричине
изменения деятельности различных физиологических систем было предложено считать
эти изменения источником эмоций. С этой точки зрения человек печален, потому
что он плачет, испытывает ярость или страх, потому что бьет другого или дрожит,
и т. д. Попытка превратить эмоции в объект, доступный объективному анализу, не
была подкреплена физиологическими опытами.
Пытаясь проверить теорию
Джемса—Ланге в эксперименте, Ч. Шеррингтон перерезал у собак блуждающие нервы и
спинной мозг, устраняя сигнализацию от внутренних органов. При этом
эмоциональные реакции на действие внешних раздражителей не исчезали. В работах
других физиологов было показано, что даже после полной изоляции головы (в
условиях перекрестного кровообращения) у кошек или собак сохраняется гамма
мимических проявлений эмоционального состояния.
Несмотря на то что теория Джемса—Ланге
низводила эмоцию до остаточных эффектов органических изменений, она явилась
толчком для многих физиологов к объективному исследованию этих изменений.
Американский физиолог У. Кеннон, первоначально сочувственно относившийся к
«периферической теории», в результате экспериментов на кошках пришел к
заключению о центральном происхождении эмоций. Он показал, что после
удаления симпатических ганглиев эмоциональные реакции сохраняются. Например,
симпатэктомированная кошка, увидев собаку, издавала угрожающие звуки,
оскаливала зубы, поднимала лапу с выпущенными когтями. Один из выводов этих
опытов был связан с признанием неспецифичности органических изменений, в
частности однонаправленности вегетативных реакций при таких различных
эмоциональных состояниях, как ярость и страх.
Как будет видно из дальнейшего
изложения, развитие физиологии показало правильность центральной теории эмоций.
Были описаны эмоциогенные структуры мозга, связанные с положительными и
отрицательными переживаниями. Вместе с тем появились новые факты,
свидетельствующие о влиянии периферических стимулов на эмоциональную сферу.
Установлена корреляция между положительными переживаниями и сокращениями мышц
лица у улыбающегося человека. Показано, что мимика изменяет кровоснабжение
мозга, т. е. может оказывать влияние на структуры, регулирующие эмоциональное
состояние. Проприоцептивная обратная связь этих рецепторов лицевой мускулатуры
также вносит вклад в его формирование. Модулирующее влияние оказывает на него и
интероцептивная сигнализация, преимущественно по афферентным системам
блуждающих нервов.
Таким образом, эмоциональное
состояние, являясь отражением активации ряда структур ЦНС, в определенной
степени зависит от висцеральных раздражителей. ,
В ряде случаев возбуждение
эмоциогенных центров вовлекает в реакцию структуры мозга, связанные с
регуляцией висцеральных процессов. Состояние последних при этом изменяется,
что, в свою очередь, оказывает модулирующее влияние на эмоциональное состояние.
Большую роль в формировании взаимодействия между центрами эмоций и интеро— и
проприоцептивными системами играет условнорефлекторная деятельность.
Наличие в структуре условного рефлекса прямых и обратных связей позволяет
понять сопряженность моторных и сенсорных функций и участие их в формировании
эмоциональных реакций.
Зависимость переживаний от
деятельности висцеральных органов наиболее рельефно проявляется при различных
патологических процессах. Нарушение кровоснабжения миокарда при спазме
коронарных сосудов сердца часто сопровождается страхом смерти. Это переживание
является рефлекторным и может быть устранено лекарственными препаратами,
расширяющими коронарные сосуды. Изменения в эмоциональной сфере часто
сопутствуют сдвигам в эндокринной системе. Больные гипотиреозом редко смеются или
плачут, у них возникает подавленность, преобладание плохого настроения.
Различные нарушения эмоциональной сферы характерны и для больных гипертиреозом.
Гуморальные раздражители могут
влиять на эмоциогенные системы мозга непосредственно и опосредованно, изменяя
состояние мотивациогенных центров. При этом положительная или отрицательная
направленность переживаний в значительной мере определяется
условнорефлекторными факторами. Например, у человека чувство голода, которое
может быть удовлетворено, окрашено положительными переживаниями, тогда как в
условиях отсутствия пищи метаболические и нервные стимулы, сигнализирующие о
потребности в ней, приводят к формированию отрицательного эмоционального
состояния.
В зависимости от
текущего возбуждения мотивационно—эмоциогенных центров мозга одни и те же
сигналы могут вызывать различные реакции. Так, периодические сокращения пустого
желудка могут инициировать или усиливать чувство голода при соответствующей
возбудимости гипоталамических отделов пищевого центра. Если же она снижена, эти
сокращения не приводят к формированию голодного побуждения. Например, в
зависимости от инструкций, полученных испытуемыми, введение им адреналина
вызывает либо гнев, либо эйфорию (повышенное благодушное настроение,
сочетающееся с беспечностью), либо не вызывает никаких эмоций.
Таким образом, хотя
периферические сигналы и участвуют в формировании эмоционального состояния,
теорию Джемса—Ланге, из которой следует, что эмоции вторичны по отношению к
органическим изменениям, нельзя считать правильной.
3.26.3. Вегетативные реакции, сопутствующие эмоциональному
состоянию
Вегетативные реакции являются важными компонентами
целостных актов, имеющих приспособительный характер, поэтому они традиционно
исследуются физиологами и психологами. Ниже перечислены некоторые из этих
реакций.
Одним из самых популярных
показателей деятельности автономной нервной системы при аффективных состояниях
у человека является кожно—гальваническая реакция (феномен Тарханова).
Она зависит от состояния потовых желез, проявляется в снижении электрического
сопротивления между двумя участками поверхности кожи. Эту реакцию используют
для определения ложных показаний, раскрытия симуляции, анализа состояния
эмоциональной сферы испытуемого.
У человека и животных аффективные
реакции отражаются также в изменениях артериального давления,
электрокардиограммы, различных показателей дыхания, температуры кожи, зрачковой
реакции, секреции слюны, пиломоторных реакций, дермографизма (изменение окраски
кожи при ее механическом штриховом раздражении), моторики желудка и кишки,
мышечного напряжения, мигания, движения глаз.
Об эмоциональной сфере судят по
содержанию катехоламинов, пептидов и глюкозы в крови, слюне и моче. Показателем
эмоционального напряжения является тремор мышц. На фоне аффекта резко меняются
различные характеристики ЭЭГ. Хотя по ЭЭГ трудно дифференцировать знак эмоции,
установлено, что отрицательные эмоциональные возбуждения характеризуются
длительным последействием и могут суммироваться; положительные —
кратковременным последействием, но устраняющим картину суммарной электрической
активности, присущей отрицательным эмоциональным состояниям. Тесно связан с
эмоциональным состоянием, преимущественно с отрицательным, гиппокампальный
Θ—ритм, однако его изменения сочетаны не только с аффективной сферой, но и
с другими сторонами деятельности мозга. Попытки связать определенную эмоцию с
возбуждением одного из отделов автономной нервной системы не увенчались
успехом. Некоторые исследователи привели доказательства связи эмоций умеренной
интенсивности, если они приятны, с парасимпатическим отделом автономной нервной
системы, если неприятны — с симпатическим.
3.26.4. Участие различных структур мозга в формировании
эмоциональных состояний
В прошлом веке было принято относить
эмоции к функции высших отделов мозга. Однако еще В. М. Бехтерев предположил,
что центром эмоций является таламус. В XX в. У. Кеннон и Ф. Бард создали
теорию, в соответствии с которой эмоционально окрашенное восприятие
определяется первоначально активизацией таламических структур, вторично
влияющих на кору больших полушарий. Д. Линдсли развил основные положения этой
теории, включив в состав эмоциогенных систем ретикулярную формацию мозга.
Согласно его «активационной теории», основанной на данных ЭЭГ, внешние и
внутренние стимулы возбуждают ретикулярную формацию ствола мозга, откуда
импульсы распространяются на гипоталамус, таламус, гиппокамп, кору больших
полушарий. Активационная теория связывает воедино эмоции, кортикальную реакцию
пробуждения, сон и бодрствование, некоторые формы патологического поведения.
Дж. Папес (1937) выдвинул иную
теорию эмоций, приписав главную роль в их формировании лимбической системе
мозга: от гиппокампа импульсы распространяются через свод в сосцевидные
тела, далее в поясную извилину, от нее — на другие области коры больших
полушарий. Значительный вклад в разработку теории эмоций внесли отечественные
физиологи (И. С. Бериташвили, П. К. Анохин, П. В. Симонов, Н. П. Бехтерева и
др.).
Наблюдения в нейрохирургической и
неврологической клиниках и опыты на животных показали, что эмоции нельзя
связать с функцией ограниченного круга мозговых структур, так как каждая из
этих структур может быть связана с положительным и с отрицательным состояниями.
В клинических случаях было
обнаружено, что электрическая стимуляция (проводимая с лечебной целью)
образований среднего мозга ведет к активации с положительной или отрицательной
эмоциональной окраской или к дезактивации, характеризующейся успокоением и
безразличием. Раздражение передней и нижней поверхностей височной доли вызывает
чувство страха; переднего и заднего отделов гипоталамуса — тревоги и ярости;
перегородки конечного мозга — наслаждения; миндалевидного тела — страха,
ярости, гнева, а в некоторых случаях и Удовольствия.
Изменения мотивационно—эмоциональной
сферы наблюдали в многочисленных опытах, проведенных на животных. Еще в конце
прошлого века Ф. Гольтц описал реакцию ярости на самые разные стимулы у собаки
с удаленной корой больших полушарий. Подобные изменения в поведении наблюдали позднее
у декортицированных и децеребрированных животных. Так как полноценное
аффективное поведение невозможно без участия в нем высших отделов мозга,
описанные особенности поведения (опущенная голова, прижатые уши, оскаленные
зубы, выпускание когтей) обозначили как «ложную ярость». Действительно,
комплекс аффективных реакций, вызываемых локальной стимуляцией подкорковых
структур, нельзя отождествлять с полноценным эмоциональным состоянием. Эти
реакции следует рассматривать лишь как отдельные элементы естественного аффекта
(рис. 3.70).
Благодаря
избирательному удалению различных структур мозга удалось определить их участие
в формировании целенаправленного поведения. Так, после разрушения латеральных
отделов гипоталамуса было обнаружено подавление мотивации к потреблению
пищи и воды, сочетающееся с изменениями в эмоциональной сфере животного.
Извращение эмоциональных реакций наблюдали у макак—резус с удаленными височными
долями, миндалевидным телом и передним гиппокампом. Особое место в этих
изменениях играло удаление миндалевидного тела. Функционально оно тесно связано с вентромедиальной
и задней областями гипоталамуса и перегородкой. Повреждение миндалевидного тела
снижает агрессивность животных, но последующее повреждение вентромедиальной области
гипоталамуса вызывает у них проявления злобности. Разрушение зоны перегородки
ведет к агрессивности, которую можно предотвратить, повредив миндалевидное
тело.
Согласно
представлениям П. Маклина, в формировании мотивационно—эмоциональных состояний
играют роль два круга: амигдалярный (миндалевидный) и септальный
(перегородочный). Первый связан с пищевыми и защитными реакциями, второй
—преимущественно с половыми.
Подкорковые
эмоциогенные структуры оказывают активирующее влияние на кору головного мозга.
Она же, в свою очередь, на основании оценки сигналов из внешней и внутренней
среды определяет формирование эмоционально окрашенных целостных поведенческих
актов.
У человека и животных существует
функциональная межполутарная асимметрия. Специализация больших полушарий
головного мозга проявляется ив эмоциональной сфере. Более эмоциогенным является
правое полушарие, играющее преимущественную роль на ранних этапах овладения
навыками. У человека положительные эмоции связаны в основном с левым
полушарием, отрицательные — с правым. Соответственно различается и оценка
афферентации от периферических рецепторов: различные психосоматические
проявления чаще связаны с левой половиной тела. Левая половина лица в большей
степени отражает отрицательные, правая — положительные эмоции. Эти различия
проявляются уже у новорожденных, в частности, в асимметрии мимики при
восприятии веществ, обладающих сладким и горьким вкусом.
Трудности в
определении локализации эмоциогенных структур мозга у животных были в
значительной степени преодолены после открытия в 1954 г. Дж. Олдсом и П.
Милнером феномена внутримозгового самораздражения
|
|
|
Рис. 3.70 Возникновение пищевых и эмоциональных реакций у животного
при поражениях ядер гипоталамуса 1 —
таламус, 2 — путь от таламуса
к гипоталамусу (перерезка), 3, 4 —
гипоталамические ядра, 5 — латеральная область гипоталамуса, 6 — супраоптическое ядро
гипоталамуса, 7 —
вентромедиальные ядра гипоталамуса; поражение вентромедиальных ядер
гипоталамуса вызывает «волчий» аппетит и ярость (А); поражение латеральной области гипоталамуса — потерю
аппетита и исхудание (Б). |

(самостимуляции). Крыса получала возможность, нажимая на педаль, замыкать
электрическую цепь и таким образом стимулировать через вживленные в мозг
электроды различные его отделы. Высокий уровень положительного подкрепления в
тех случаях, когда электрод находился в положительной эмоциогенной структуре
(«зоны награды», «центры удовольствия», «система поощрения»), приводил к
выработке инструментального условного рефлекса в течение нескольких минут.
«Зоны награды» созревают очень рано
и уже в первые дни после рождения включаются в регуляцию поведения. Система
отрицательного подкрепления («зоны наказания») также готова к функционированию на
самых ранних этапах постнатальной жизни. Если электрод вживлен в «зону
наказания», животное, замкнув один раз электрическую цепь, в дальнейшем
избегает нажатий на педаль.
Таким образом, по стремлению
животного к максимизации и минимизации электростимуляции данной «точки» можно
отнести ее к системам положительного или отрицательного подкрепления.
3.26.5. Эмоциогенные системы мозга
У крысы 35% клеточного пространства
мозга связано с положительной эмоциогенной системой, только 5% — с
отрицательной, 60% — с отсутствием побуждения к повторению или избеганию
стимуляции. Принципиальная организация эмоциогенных систем мозга оказалась
весьма сходной у представителей разных отрядов млекопитающих. На рис. 3.71
представлена топография эмоциогенных систем мозга.
В процессе обучения животные выбирают силу тока и ритм
нажатий на педаль, ведущие к наибольшему «вознаграждающему» эффекту. Как и
естественное аффективное поведение, внутримозговая самостимуляция
сопровождается комплексом вегетативных реакций симпато—парасимпатической
природы.
|
|
|
Рис.
3.71.Топография
эмоциогенных зон (заштриховано) мозга у крысы (А), кошки (Б), обезьяны (В) и
человека (Г) |
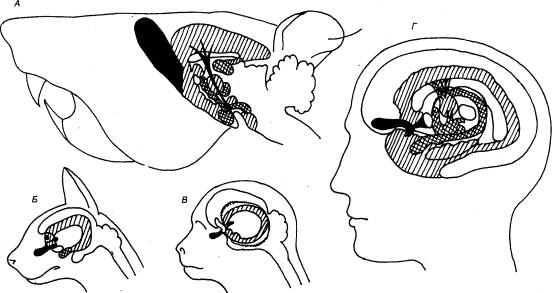
«Зоны награды» находятся
в тесном соседстве с мотивациогенными структурами мозга. В некоторых случаях
слабое принудительное раздражение определенной «точки» вначале вызывает
потребление пищи, воды или другое поведение, направленное на удовлетворение
биологической потребности. При увеличении силы тока животное переходит к
самостимуляции. В других случаях можно выделить «точки», изолированно связанные
только с определенной мотивацией или только с реакцией самораздражения.
Следовательно, мотивациогенные и эмоциогенные структуры представляют собой хотя
и функционально взаимозависимые, но дискретные нейроанатомические системы.
Опыты, проведенные на щенках,
начиная с периода новорожденности, показали, что на разных этапах онтогенеза наблюдается
различное взаимодействие мотивациогенных и эмоциогенных систем мозга.
Свойственное взрослому животному единство мотивационно—эмоционального поведения
является результатом формирования сложных условнорефлекторных стереотипов,
отражающих опыт существования в определенной внешней среде, т. е. наиболее
целесообразных для приспособления к конкретной обстановке.
Пластичность поведения хорошо прослеживается в
экспериментах с телестимуляцией различных мозговых структур (X. Дельгадо). Раздражение определенных
участков таламуса или центрального серого вещества вызывало у вожака колонии
обезьян повышенную агрессивную реакцию, однако это проявлялось лишь по
отношению к сопернику; он никогда не нападал на маленькую самку, свою любимую
подругу. Показано, что в зависимости от ситуации стимуляция одной и той же
«точки» может вызывать различные целенаправленные действия (пищевые, питьевые,
половые, защитные, агрессивные реакции) с присущей им эмоциональной окраской. В
соответствии с подобной пластичностью мотивационно—эмоциональных реакций
телеметрическая стимуляция одной и той же «точки» мозга вызывает самые разные
формы поведения в зависимости от того, исследуется ли оно в лабораторных
условиях или в экологически адекватной среде.
Эмоциональные состояния строятся на
основе определенных нейрохимических механизмов. Фармакологический анализ
показал, что настроение у человека и самостимуляция «зон награды» у животных
зависят от нейромедиаторного баланса в ЦНС. Однако попытка связать определенное
эмоциональное состояние с уровнем какого—либо медиатора потерпела неудачу.
«Точки» мозга, в которых можно
вызывать самораздражение, связаны с дофаминергическими и норадренергическими
системами, снижение их активности подавляет самостимуляцию «зон награды».
Возможно, однако, что положительное эмоциональное возбуждение определяется
норадреналином, тогда как дофамин лишь активирует систему побуждения к
самораздражению.
По другим предположениям,
подкрепляющая функция эмоций зависит от дофамин— и серотонинергических систем,
а мотивационные реакции — от адренергической.
Несомненно доказано участие в
формировании положительного эмоционального состояния эндогенных опиатов. В свою
очередь, эндорфины модулируют состояние серотонинергической системы. По мнению
некоторых исследователей, серотонинергическая и холинергическая системы
обслуживают отрицательные эмоции. Существуют, однако, сведения о подавлении
агрессивных тенденций серотонином.
Таким образом, можно предполагать,
что в реализации различных эмоциональных состояний участвуют все
нейромедиаторные системы мозга.
3.26.6. Влияние эмоциональных состояний на научение и память
Связь мотивациогенных и эмоциогенных
систем мозга проявляется в существенной роли, которую играет уровень
мотивационно—эмоционального возбуждения в процессах научения. Оно затруднено
при низком уровне мотивации и подкрепления. Высокий уровень потребности
формирует выраженное эмоциональное возбуждение. Оно наиболее четко проявляется
на первой стадии выработки условного рефлекса — стадии генерализации, по мере
его упрочения постепенно снижается. Как можно видеть по ряду вегетативных
проявлений (например, частоте сердцебиений), даже оборонительные условные
рефлексы после их упрочения реализуются автоматически без каких—либо проявлений
эмоционального возбуждения.
Оборонительные условные рефлексы,
связанные с отрицательным подкреплением, формируются значительно быстрее и
длительнее сохраняются, чем условные рефлексы, вырабатываемые при положительном
подкреплении (например, пищедобывательные). Так, условнорефлекторные вкусовые
аверсии — отказ от пищи, потребление которой вызвало отравление, —
вырабатываются при однократном сочетании вкусовых сигналов с последущим
дискомфортным состоянием, причем этот рефлекс может сохраняться в течение
многих месяцев, а иногда и лет.
Принято считать, что
запоминание и длительность сохранения памятного следа пропорциональны
эмоциональному напряжению. Однако, как показали эксперименты на добровольцах,
низкий и высокий его уровни отрицательно сказываются на обучении и процессах
памяти. Наиболее благоприятным для них оказался умеренный уровень
эмоционального напряжения, соответствующий умеренной активации ЦНС. Наблюдения
на школьниках разного возраста показали, что в I—III, IX и Х классах положительное
подкрепление улучшало, а в IV—VIII — ухудшало запоминание. Полагают, что
разнонаправленные влияния эмоционально—положительной и отрицательной систем на
мнестические процессы данных возрастных групп определяются гормональными
факторами, соответствующими .этим периодам развития.
В опытах на животных
было показано, что условнорефлекторные реакции, подкрепляемые
непосредственной электрической стимуляцией эмоциогенных структур мозга,
формируются очень быстро. При этом можно переделать значимость исходно
отрицательных раздражителей. Так, у собак вырабатывали положительную реакцию на
вкус горького раствора, сочетаемого со стимуляцией «зон награды». Этот же
механизм, по—видимому, лежит в основе условнорефлекторного формирования
предпочтения человеком веществ с исходно индифферентными или отрицательными
вкусовыми свойствами, если они оказывают положительный эмоциональный эффект.
Привычка потреблять эти вещества может быть чрезвычайно сильной, а в некоторых
случаях перейти в патологическое влечение. Доказано, что начальные стадии
формирования наркомании определяются эйфорией, связанной с активацией «зон
награды».
Влияние эмоциональных
состояний на научение и память определяется также модулирующим действием
подкрепляющих систем мозга на сенсорные процессы (зрительная, слуховая,
вкусовая, болевая и другие системы). В частности, стимуляция «зон награды»
изменяет электрические характеристики нейронов ряда анализаторов на разных
уровнях мозга, т. е. эмоциогенные системы мозга оказывают модулирующее влияние
на обработку сенсорной информации.
Влияния, оказываемые
эмоциональными состояниями на сенсорные процессы, научение, память,
характеризуются разной степенью напряженности.
В ряде случаев преимущественно
обращают внимание на аффекты, крайние эмоциональные состояния,
характеризующиеся выраженными субъективными и объективными показателями. Однако
эмоциональная окраска присуща любой деятельности и не обязательно проявляется
как ярко окрашенное переживание. Состояния общего комфорта или дискомфорта,
отражающие баланс между уровнем активности положительных и отрицательных
эмоциогенных систем, являются основой организации целостных поведенческих
актов.
Наряду с исследованием
физиологических механизмов аффективных состояний было обращено внимание на
разработку общетеоретических представлений о роли эмоций в организации
поведения. Согласно биологической теории П. К. Анохина, эмоции возникли в
процессе эволюции как средство быстрой оценки потребностей и их удовлетворения
в соответствии с состоянием внешней ситуации. Если параметры достигнутого
эффекта соответствуют свойствам акцептора результатов действия, возникает
положительная эмоция, если не соответствуют — отрицательная.
Потребностно—информационная теория П. В. Симонова рассматривает эмоцию
как отражение мозгом человека и высших животных качества и величины определённой
потребности и вероятности ее удовлетворения. Регуляторная роль эмоций
проявляется в их отражательно—оценочной, переключающей, подкрепляющей,
компенсаторной (замещающей) функциях. Подобный подход к рассмотрению эмоций
сближает физиологические, психологические и философские аспекты изучения этого
важнейшего феномена в поведении человека и животных.
Другая сторона исследования эмоций
связана с разработкой вопросов их нарушения. Ослабление деятельности
систем положительного подкрепления или усиление деятельности системы
отрицательного подкрепления приводит к гипотимии (от греч. hypo — понижение и thymos — настроение, чувство) —
депрессивному состоянию с проявлением тревоги, страха, апатии. При этом
происходит ослабление мотивационных процессов, снижение массы тела, нарушение
работы внутренних органов.
При гипертимии наблюдаются
повышенное настроение, усиление двигательной активности, легкая отвлекаемость.
Нередко фазы гипотимии и гипертимии перемежаются. У человека описаны и другие
разновидности нарушения эмоциональной сферы, причины которых различны. Как
правило, в основе глубоких нарушений эмоциональной сферы лежат генетические
факторы. Обычно гипо— и гипертимия сопровождаются значительными отклонениями в
балансе нейромедиаторов. Так, при маниакальных и депрессивных состояниях
изменяется синтез дофамина, норадреналина и серотонина в разных отделах мозга.
Ряд симптомов этих заболеваний, в частности, тревожность и депрессию, можно
воспроизвести на животных с помощью введения нейротоксинов или фармакологических
веществ, подавляющих синтез норадреналина, серотонина и опиатов.
При взаимодействии человека и животных с внешней средой
возникают ситуации, характеризующиеся конфликтом между потребностями и возможностями
их удовлетворения. Они ведут к формированию состояния напряжения — эмоционального
стресса, приспособительное значение которого проявляется в мобилизации
защитных сил, направленных на преодоление конфликта. Невозможность его
разрешения приводит к формированию длительного застойного эмоционального
возбуждения, проявляющегося в нарушениях мотивационно—эмоциональной сферы и в
различных соматических заболеваниях.
Эмоциональный стресс может привести
к развитию ишемической болезни сердца, гипертензии, язвообразованию, дисфункции
эндокринной системы. При этом наблюдаются глубокие изменения баланса
нейромедиаторов и нейропептидов в ЦНС. У человека такие нарушения связаны
преимущественно с социальными конфликтами. Некоторые из этих проявлений могут
быть смоделированы на животных. Так, у вожака стада обезьян, изолированного, но
имеющего возможность наблюдать последующие иерархические изменения во
взаимоотношениях ранее подчинявшихся ему животных, развивается артериальная
гипертензия, а в некоторых случаях и инфаркт миокарда.
Степень устойчивости животных к действию стрессорных
факторов различна и может быть установлена в специальных экспериментах. Так,
менее устойчивые к стрессу животные отвечают на стимуляцию отрицательных
эмоциогенных зон вентромедиального гипоталамуса преимущественно прессорными
сосудистыми реакциями, а более устойчивые — прессорно—депрессорными. Снижение
эмоционального напряжения и, соответственно, предотвращение висцеральных
нарушений достигается стимуляцией «зон награды» или введением фармакологических
препаратов, нормализующих баланс медиаторов в ЦНС.
Исследование различных проявлений
эмоционального стресса оказалось полезным для разработки проблемы неврозов —
группы обратимых функциональных расстройств психогенной природы. В отечественной
литературе принято выделять три формы невроза: истерический, навязчивых
состояний и неврастению. Возникновение невроза и его форма определяются
взаимодействием психотравмирующей ситуации с исходными характеристиками
личности. В ряде случаев невротическое состояние развивается на фоне
недостаточности структур лимбико—ретикулярного комплекса. Эмоциональные
нарушения занимают в симптоматике неврозов одно из ведущих мест.
И. П. Павловым было введено понятие экспериментальные
неврозы. — функциональные нарушения (срыв) высшей/нервной деятельности в
результате перенапряжения основных нервных процессов. Учение И. П. Павлова об
экспериментальных неврозах оказалось плодотворным для понимания ряда сторон
патологии психической деятельности у человека. Однако нельзя отождествлять
сложную клиническую картину невроза у человека с нарушениями поведения у
животных, на которых можно лишь моделировать определенные симптомы заболевания.
Тенденция к отождествлению большей части функциональных нарушений поведения
животных с экспериментальными неврозами создала положение, которое затрудняет
прямое сопоставление клинических и экспериментальных данных. Вместе с тем
возможность исследовать у животных отдельные проявления невроза у человека, а
также проводить их фармакологический анализ, приносит существенную пользу
неврологической клинике.
Основными приемами выработки экспериментального невроза И.
П. Павловым и его сотрудниками было создание конфликтной ситуации при
столкновении разнонаправленных мотиваций, например, пищевой и оборонительной,
или же при необходимости решения трудной задачи (дифференцирование близких по
параметрам условных сигналов и т. п.).
Массовое проявление невротических реакций у собак,
находившихся в виварии, затопленном во время ленинградского наводнения 1924 г.,
позволило И. П. Павлову сделать ряд важных обобщающих выводов о закономерностях
развития патологии высшей нервной деятельности. Он постулировал зависимость
невротизации от силы, подвижности и уравновешенности корковых
процессов возбуждения и торможения. В соответствии с ними была предложена
классификация темпераментов у человека.
Эксперименты, проведенные на
собаках, показали, что неврозы легче возникают у животных слабого и сильного
неуравновешенного типов высшей нервной деятельности. Анализируя происхождение
истерии, И. П. Павлов рассматривал ее как следствие слабости нервной системы.
Современные методы исследования усложнили эту схему.
Оказалось, что при экспериментальном неврозе функциональные изменения возникают
прежде всего в лобных отделах новой коры, лимбических структурах и ретикулярной
формации среднего мозга. Нарушается баланс нейромедиаторов в ЦНС и в крови, в
частности, между катехоламинами и ацетилхолином. Хотя неврозы и рассматривают
как функциональные заболевания, выяснилось, что они сопровождаются развитием
реактивных и дегенеративных процессов в различных отделах мозга. Показана роль
информационных факторов в развитии патологии мотивационно—эмоционального
поведения. Большую роль при этом играют нарушения памяти.
Изучение патологии высшей нервной деятельности подтвердило
роль обратной связи от соматических и висцеральных систем в поддержании
отрицательного эмоционального состояния. Были приведены факты,
свидетельствующие в пользу существования эмоциональной памяти
(«аффективной памяти», «памяти чувств»), на которую впервые обратил внимание в
конце прошлого века психиатр С. С. Корсакова, а И. С. Бериташвили выделил ее
как особую форму памяти.
Она особенно четко проявляется при неврозах, в основе
которых лежат инертные очаги возбуждения (травматические неврозы,
патологические влечения — алкоголизм, наркомания).
Эмоциональные условные реакции могут усиливаться или
тормозиться внешними раздражителями, преимущественно ситуационными. Так, уже
упоминавшиеся вкусовые аверсии могут быть резко ослаблены в привычной
обстановке и в полной мере проявиться в непривычной. У человека и животных
отрицательные эмоциональные реакции со всем комплексом вегетативных проявлений
могут воспроизводиться при столкновении с ситуацией, сопровождавшейся
травматизирующими факторами, даже если со времени их воздействия прошло много
лет.
Эмоциональное состояние определяется генотипическими
характеристиками и условиями, в которых проходило развитие животных. Особо
важную роль при этом играют ранние воздействия. Так, изоляция от сверстников
или от взрослых особей в раннем онтогенезе повышает неустойчивость к
невротизирующим стимулам. Обогащенная или обедненная раздражителями внешняя
среда сказывается на морфофункциональном развитии мозга, в частности, его
эмоциогенных систем. Эти данные, полученные на животных, позволяют разработать
рекомендации для создания условий воспитания детей, начиная с периода
новорожденности, способствующих оптимальному развитию их
мотивационно—эмоциональной сферы.
3.27. ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР
Из пищеварительного тракта в кровь
поступают продукты расщепления пищи, и некоторые из них в случае проникновения
в мозг могли бы нарушить его функции. Например, переваривание белков приводит к
повышению уровня в плазме крови аминокислот глицина и глутамата, которые в
мозге играют роль медиаторов. Высокая двигательная активность также нарушает
постоянство внутренней среды организма. Поведение низших животных, не имеющих
гематоэнцефалического барьера (ГЭБ), сильно зависит от состава пищи и
равномерности ее поступления в организм, что привязывает каждый вид к
конкретному источнику питания и делает невозможными активные перемещения в
пространстве для эффективного поиска пищи или половых партнеров.
Возникновение в ходе эволюции
способности к зоосоциальному поведению и простейшим формам научения было
связано с усложнением структуры и функций мозга, что потребовало поддержания в
нем более строгого постоянства внутренней среды. Поскольку достичь такой
степени гомеостатирования во всем организме невозможно, у высокоорганизованных
животных вокруг ЦНС выделилось особое пространство, окруженное барьером,
непроницаемым для большей части веществ, содержащихся в крови (гемолимфе) и
тканевой жидкости. К двум хорошо известным пространствам — внутриклеточному и
внеклеточному — добавилось третье пространство, содержащее
цереброспинальную (спинномозговую) жидкость, или ликвор. Так эволюция сделала
шаг сопоставимый по значению с появлением внутренней среды организма.
Создание особой внутренней среды
мозга, во—первых, защитило его нейроны от колебаний внутренней среды организма,
связанных с потреблением пищи и двигательной активностью, и обеспечило
стандартные условия для интегративной деятельности нейронов и синаптической
передачи; во—вторых, позволило нейронам мозга при передаче друг другу сообщений
гуморальным путем экономно использовать запасы нейросекретов и уменьшило
искажение передаваемой информации.
Потребность в барьере
была настолько сильной, что он независимо возник не менее чем в трех ветвях
эволюции, выделившихся 500—600 млн. лет назад, и имеется у всех таксонов,
способных к сложному поведению: у позвоночных головоногих моллюсков и
членистоногих (у насекомых, некоторых паукообразных и отчасти у ракообразных).
Структура барьера. Мысль о существовании барьера между
кровью и мозгом впервые высказал Пауль Эрлих в 1885 г. К этому времени
накопились данные о том, что красители (например, трипановый синий) после
введения их в кровь не обнаруживаются в мозге. В 1913 г. Голдманн показал, что
если краситель вводить не в вену, а в ликвор, то мозг окрашивается. Тем не
менее долгое время существовала и противоположная точка зрения: барьера нет,
красители не проходят в мозг потому, что мозг не содержит соединительной ткани
т. е. для красителя в мозге просто нет места. Только в 60—е годы мнение о
существовании ГЭБ стало общепринятым.
Термин «гистогематические барьеры» предложила наша соотечественница Л. С. Штерн в 1929 г. Сейчас он понимается как общее название для барьеров: гематоэнцефалического (ГЭБ),
—ликворного (ГЛБ), —неврального, —офталъмического, —тестикулярного,
—лабиринтного, —плеврального, —синовиального
и др. В 1933 г. Вальтер и Шпац
ввели понятие «гематоэнцефалический
барьер».
Как известно, капилляры мозга принципиально отличаются от
капилляров всех других областей тела отсутствием в их стенке водных каналов,
пор, или фенестр. Под электронным микроскопом отчетливо видно, что
эндотелиоциты мозговых капилляров соединены между собой так называемыми плотными
контактами, которые не позволяют водорастворимым веществам проходить из
крови в головной мозг (и обратно). На электронограммах также видны и отростки астроцитов,
охватывающие всю (или почти всю) наружную поверхность мозговых капилляров (рис.
3.72).
Эти отростки не составляют механического препятствия
проникновению веществ в мозг, как думали прежде, но они секретируют некие пока
недостаточно изученные, вещества, повышающие плотность контактов между
|
|
|
Рис. 3.72 Ножки отростков астроцитов (А)
окружают мозговой капилляр (К) |
|
|
|
Рис. 3.73 «Энзимный барьер» МАО — моноаминооксидаза, КОМТ —
катехол—0—метил—трансфераза, ЭП — эпендима, ЭН — эндотелий. |
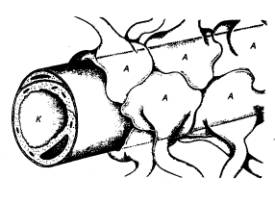
эндотелиоцитами. Известно, что по
крайней мере часть этих веществ имеет молекулярную массу менее 4 кДа и,
следовательно, не относится к белкам.
В середине 60—х годов было открыто существование «энзимного
барьера», т. е. ферментов в тканях, лежащих между просветом капилляров и
нейронами мозга. Эти ферменты расщепляют вещества, которые в противном случае
могли бы проникнуть из крови в мозг. Примерами таких «барьерных» энзимов могут
служить катехол—0—метилтрансфераза (КОМТ), моноаминоксидаза (МАО),
тирозингидроксилаза и аминопеппгидаза. Энзимный барьер в большей
степени препятствует проникновению в мозг моноаминов (потенциальных
нейротрансмиттеров), чем их непосредственных предшественников (рис. 3.73).
Феномен ГЭБ пытались объяснить еще и
тем, что введенный в кровоток краситель связывается с белками плазмы и это
делает его неспособным проникнуть сквозь барьер. Однако связывание с белками
плазмы не препятствует окрашиванию всех прочих (немозговых) тканей, поэтому
сегодня фактору связывания придается небольшое значение.
Итак, ГЭБ состоит, по меньшей мере,
из трех главных компонентов: (1) плотных контактов в эндотелии капилляров; (2)
веществ, секретируемых отростками астроцитов и поддерживающих функцию плотных
контактов; и (3) «барьерных энзимов».
Проницаемость барьера и транспортные системы. В 1902 г. Эрлих заметил, что
красители, поглощаемые мозгом, поглощаются и жировой тканью тоже. Значение
этого факта было оценено только в 70—е годы, после создания высокоточного
метода количественной оценки проницаемости ГЭБ.
Для жирорастворимых веществ ГЭБ барьером вообще не является: они
простой диффузией легко проходят сквозь эндотелиоциты: сначала растворяются в
липофильных участках люминальной (т. е. обращенной в сторону просвета
сосуда) мембраны, затем диффундируют через всю толщу цитоплазмы и, наконец,
растворяясь в аблюминальной (расположенной с другой, наружной по
отношению к просвету сосуда стороны) мембране, выходят из эндотелиоцита в ткань
мозга (рис. 3.74, путь 1А).
Никакие живые клетки не являются препятствием для веществ,
обладающих высокой липофильностью (или, что то же самое, гидрофобностью).
Наиболее высокой «проходимостью» обладает углекислый газ, легко
|
|
|
Рис. 3.74 Пути проникновения веществ из крови в мозг ГЭБ — гематоэнцефалический барьер;
ЦВО — циркумвентрикулярные органы; ВСС — внутрижелудочковое сосудистое
сплетение; ЦСЖ — цереброспинальная жидкость (ликвор). 1 — сквозь нефенестрированный эпителии: А — простой диффузией;
Б — активным транспортом; 2 —
через фенестры эндотелия капилляров в ЦВО и далее: А — внутриаксональным
транспортом; Б — сквозь эпендиму простой диффузией в ликвор; В — вдоль
наружной границы эпендимы желудочков до ее участков, где нет плотных
контактов (гипотетический путь); 3 — сквозь стенки сосудистого сплетения в
ликвор (простой диффузией); 4 —
со слизистой оболочки полости носа (конкретный путь неясен); 5 —
внутриаксональным транспортом (например, по чувствительным нейронам). Для
жирорастворимых веществ наибольшее значение имеют пути 1А, 2Б и 3, для
водорастворимых — 2А, 2В, 4 и 5. Для водорастворимых веществ, нужных мозгу в
значительных количествах (глюкоза, аминокислоты и др.) существуют системы
переносчиков. |
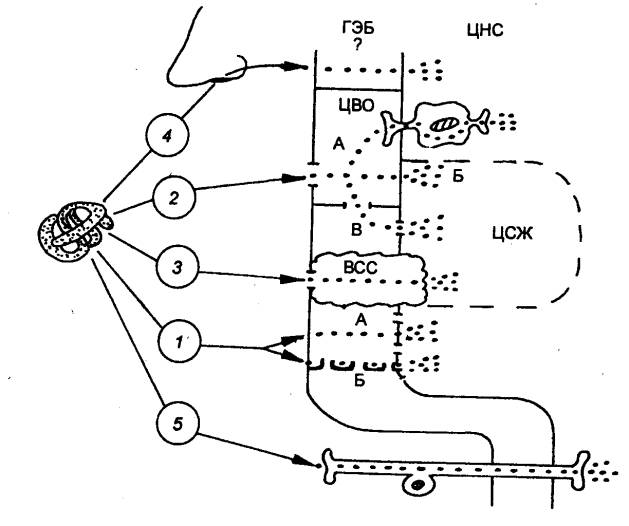
проникающий
из крови в нейроны
дыхательного центра. Довольно высокой растворимостью, соответственно,
«проходимостью» обладают героин, алкоголь и никотин, что объясняет их быстрое (после
попадания в кровь) действие на ЦНС. Следовательно, липофильность — главное из
качеств, определяющих возможность простои диффузии вещества сквозь ГЭБ. На «проходимость» вещества
диффузией также влияют степень его ионизации, молекулярная масса и некоторые
другие факторы.
Мозг нуждается в некоторых веществах, не обладающих
способностью растворяться в жирах, например, в глюкозе — для обеспечения своих
энергетических потребностей, и в аминокислотах — для синтеза белков. Поскольку
гидрофильные вещества простой диффузией пройти в мозг не могут, для их доставки
существуют транспортные системы со специфическими белками—переносчиками (рис. 3.74, путь 1Б). Сейчас известны
транспортные системы для: (а) D—глюкозьг; (б) крупных нейтральных аминокислот;
(в) основных и кислых аминокислот; (г) электролитов (K+, Mg2+, Ca2+, I— и др.); (д)
водорастворимых витаминов; (е) нуклеозидов и др. Все эти системы обладают
общими свойствами: селективностью, стереоспецифичностью, конкурентным
ингибированием и насыщаемостью,
Мелкие нейтральные аминокислоты не транспортируются, так как
мозг синтезирует их сам. Транспортная система, переносящая глюкозу, не требует
для своей работы присутствия инсулина. Необычно то, что в ГЭБ инсулин регулирует транспорт не
глюкозы, а аминокислот.
«Дефекты» в
барьере и пути в обход барьера. В головном мозгу есть области, в которые, в
отличие от основной его массы, краситель легко проходит из крови. Впервые это
обнаружил Шульманн в 1912г.: трипановый синий, введенный мышам внутривенно,
проникал в нейрогипофиз. Количество областей, обладающих такими особенностями,
постепенно росло, и сейчас включает:
нейрогипофиз,
медиальное возвышение, самое заднее поле, субфорникальный орган, шишковидное
тело, зрительное углубление, сосудистое сплетение и сосудистый орган конечной
пластинки. Поскольку все эти структуры расположены по периферии желудочковой
системы, было предложено называть их циркумвентрикулярными органами (рис.
3.75). Только один из них не обладает названными особенностями: в
субкомиссуральном органе эндотелиоциты соединены плотными контактами.
|
|
|
Рис. 3.75 Циркумвентрикулярные органы Э — шишковидное тело (эпифиз), ВСС — сосудистое сплетение,
СКО — субкомиссуральный орган, СФО — субфорникальный орган, MB — медиальное возвышение, СОКП — сосудистый
орган конечной пластинки, НГ — нейрогипофиз, СЗП — самое заднее поле. |
|
|
|
Рис. 3.76 Принципиальное различие
устройства гематоликворного (А) и гематоэнцефалического (Б) барьеров Плотные
контакты (ПК) соединяют либо клетки эндотелия капилляров (К), либо клетки
эпендимы (Э), выстилающей стенки желудочков мозга. ИП — интерстициальное
пространство; Н — нейрон; Г — глия; Ф — поры (фенестры); ЦСЖ —
цереброспинальная жидкость (ликвор); S — площадь барьера. Стрелками
показано проникновение водорастворимых веществ. |
В циркумвентрикулярных органах переносимые кровью вещества относительно
свободно покидают просвет капилляров (рис. 3.74., путь 2) и доходят до наружной
границы желудочков мозга, где их задерживают плотные контакты, которыми соединены
клетки выстилающей желудочки эпендимы. Эту особую организацию барьера в
циркумвентрикулярных органах часто ошибочно принимают за «дефекты» в барьере. В
действительности, барьер в этих областях не менее эффективен, только
разделительную функцию выполняет не эндотелий капилляров, а эпендима желудочков
мозга. Таким образом, барьер здесь просто отнесен чуть далее вглубь мозговой
ткани и носит название гематоликворного (ГЛБ). Площадь ГЛБ составляет
1/5000 общей площади ГЭБ (рис. 3.76).
Полагают, что некоторая часть молекул веществ, оказавшихся в
циркумвентрикулярных органах, может проникнуть в собственно мозг:
— через нервные окончания и далее внутриаксональным
транспортом (см. рис. 3.74, 2А);
— сквозь клетки эпендимы — в ликвор,
в соответствии с растворимостью данного вещества в липидах (рис. 3.74, 2Б);
— перемещением вдоль наружной границы эпендимы до тех участков,
где между ее клетками есть водные каналы, и по ним — в ликвор (рис. 3.74, 2В).
Некоторая часть сосудов мозга выпячивается в желудочковую систему
и образует в ней сосудистое сплетение — орган, продуцирующий ликвор.
Капилляры этих сплетений имеют фенестры, сквозь которые вещества легко покидают
кровь, но в дальнейшем задерживаются хориоидным эпителием, составляющим единое
целое с остальной эпендимой. Дальнейшее продвижение (в ликвор) возможно только
простой диффузией, в соответствии с липофильностью конкретного вещества (рис.
3.74, путь 3).
Возможно,
единственный путь, по которому водорастворимые вещества в значительных
количествах без помощи специальных транспортных систем попадают в мозг,
начинается на слизистой оболочке полости носа (рис. 3.74, путь 4). Пероксидаза
хрена (белок с молекулярной массой 40 кДа), введенная интраназально, за
несколько минут достигает пиальной поверхности обонятельных луковиц. Водные
растворы нейропептидов вызывают ЦНС—зависимые эффекты (например,
интраназальное введение человеку вазопрессина приводит
|
|
|
Рис. 3.77 Пути воздействия веществ на мозг без их
проникновения сквозь гематоэнцефалический барьер 1—3 — связывание веществ со специфическими рецепторами в
циркумвентрикулярных органах приводит к: увеличению проницаемости ГЭБ для
другого вещества (1)—образованию в циркумвентрикулярных органах
жирорастворимых веществ, проникающих далее сквозь эпендиму желудочков простои
диффузией (2А); образованию в самом мозгу гипотетических водорастворимых
веществ—посредников (2Б); передаче возбуждения в виде потенциалов действия
(3); 4 — связывание с пресинаптическими рецепторами эфферентов имитирует
центральное действие; 5 — воздействие на периферические органы через
возбуждение центральных афферентов изменяет состояние ЦНС. Остальные
обозначения, что и на рис. 3.74 |
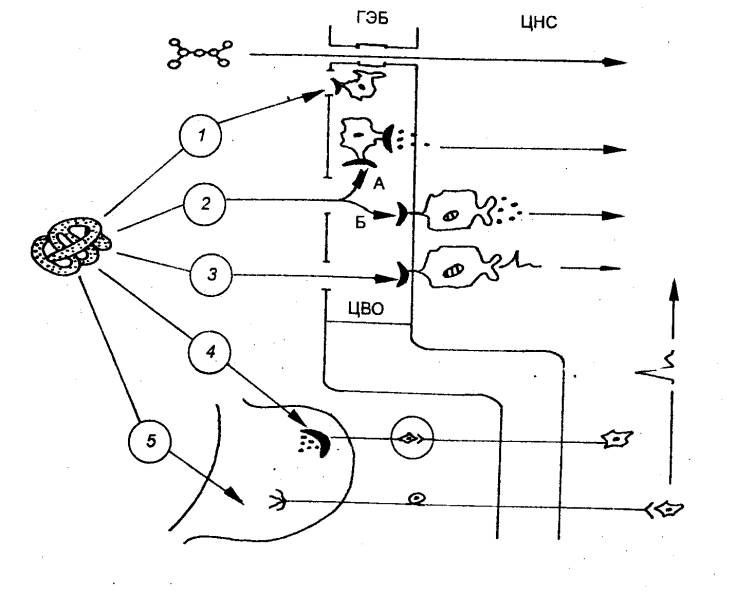
к отчетливому повышению настроения).
В этом случае диффузия происходит либо внутри аксонов обонятельных нейронов,
либо через подслизистый слой связанный с подпаутинным пространством в области
обонятельных луковиц. Нарушение целостности слизистой оболочки (например, при
рините) способствует проникновению веществ в мозг именно этим путем.
Гидрофильные
молекулы из крови могут попадать в мозг внутриаксональным транспортом,
например, по волокнам чувствительных нейронов (рис. 3.74, путь 5)
Рецепторы
наружной поверхности барьера. Есть несколько механизмов посредством которых
водорастворимые вещества, находящиеся в плазме крови могут воздействовать на
мозг без проникновения в него (рис. 3.77), и часть этих механизмов связана с
существованием на «внешней» стороне барьера специфических рецепторов к многочисленным
гормонам белково—пептидной природы
В 50—е годы было
показано, что если разрушить отдельные циркумвентрикулярные органы, то
некоторые гормоны, переносимые плазмой крови теряют способность действовать на
мозг. Позднее были открыты расположенные в циркумвентрикулярных органах
рецепторы к соматостатину, кортикотропину (адренокортикотропному гормону),
пролактину, вазопрессину, окситоцину паратирину (паратгормону), кальцитонину,
инсулину и инсулиноподобным факторам роста, эндотелину, панкреатическому полипептиду,
ангиотензину II атриальному и мозговому натрийуретическим пептидам,
брадикинину, вазоактивному интестинальному полипептиду, субстанции Р и
нейропептидам У и YY
Возбуждение
«внешних» рецепторов молекулами одного вещества может увеличить проницаемость
барьера для другого находящегося в плазме крови вещества (рис. 3.77, путь 1).
Это может выражаться в стимулировании работы
одной из систем активного транспорта (например, внутривенное введение
вазопрессина увеличивает транспорт в мозг лейцина, некоторые гормоны регулируют
проникновение в мозг моноаминов) или в снижении сопротивления барьера простой
диффузии веществ (так, вазопрессин увеличивает проницаемость ГЭБ для молекул
воды). Взаимодействие лиганда с рецептором
на наружной стороне барьера может также приводить к образованию
веществ—посредников, способных воздействовать непосредственно на нейроны мозга
(рис. 3.77, путь 2). Место синтеза таких посредников определяется их
способностью проникать сквозь барьер. Жирорастворимые вещества—посредники синтезируются
на внешней стороне барьера, как правило, в циркумвентрикулярных органах (см.
рис. 3.77, путь 2А). Так, образующиеся в периферических тканях интерлейкины —
полипептиды с большой молекулярной массой (примерно от 8 до 26 кДа) и низкой
растворимостью в жирах, попадая в сосудистый орган конечной пластинки,
стимулируют образование в нем липофильных молекул простагландинов, которые
проникают в мозг и, воздействуя на центры терморегуляции, вызывают повышение
температуры тела — характерное проявление лихорадки.
Водорастворимые вещества—посредники,
не обладающие способностью проникать сквозь ГЭБ, могут синтезироваться и
высвобождаться только «за барьером», в самом мозге (рис. 3.77, путь 2Б). Иногда
в качестве посредника выступает то же самое вещество, которое воздействовало на
«внешние» рецепторы. Так, в ответ на поступление пищи в кишку расположенные в
ее слизистой оболочке эндокринные клетки высвобождают в кровь холецистокинин
(ХЦК). Одновременно возрастает и концентрация ХЦК в мозге. Однако ХЦК, как
и большинство других пептидов, не обладает сколько—нибудь заметной
растворимостью в жирах, поэтому простой диффузией пройти в мозг он не может.
Специальной транспортной системы для ХЦК в барьере также не обнаружено.
Полагают, что молекулы ХЦК, не проникая в ЦНС, связываются с
холецистокининовыми рецепторами на наружной стороне ГЭБ и активируют
неизвестный пока механизм, способствующий высвобождению «по ту сторону барьера»
таких же молекул ХЦК, но синтезированных в самом мозге. Этот механизм играет
важную роль и в возникновении такого чувства, как насыщение.
Предполагают также, что
чувствительные окончания некоторых нейронов мозга выходят на внешнюю сторону
ГЭБ и через них возбуждение от перечисленных выше рецепторов может передаваться
в мозг виде потенциалов действия (рис. 3.77, путь 3).
Таким образом, с помощью
циркумвентрикулярных органов мозг отвечает на колебания концентраций в крови
многочисленных биологически активных веществ. При этом циркумвентрикулярные
органы могут быть столь же непроницаемы, как и другие части ГЭБ.
Вещества, циркулирующие с кровью,
могут воздействовать на ЦНС и опосредованно, через действие на периферические
органы (рис. 3.77, путь 4). Например, пептидный гормон бомбезин
связывается со специфическими рецепторами в гладких мышцах пилорической части
желудка, что приводит к повышению мышечного тонуса привратника и способствует
задержке пищи в желудке. Импульсация от механорецепторов стенки заполненного
пищей желудка по чувствительным волокнам блуждающего нерва передается в ЦНС, в
результате чего частично подавляется пищевое поведение. Так воздействие
вещества на периферический орган вызывает поведенческий (т. е. центральный)
эффект.
Гидрофильные вещества, растворенные
в плазме крови, могут не только действовать на ЦНС, но и имитировать такое действие,
усиливая или ослабляя высвобождение медиаторов из периферических нервных
окончаний (рис. 3.77, путь 5). Так, тиролиберин и ангиотензин II,
связываясь с пресинаптическими рецепторами симпатических нервных окончаний,
облегчают высвобождение ими норадреналина, имитируя влияние на тонус
симпатических нейронов спинного мозга.
Пути выхода веществ из мозга в
кровь. Специальные
транспортные системы выносят из мозга в кровь метаболиты самого мозга (их
накопление могло бы
|
|
|
Рис. 3.78 Неизбирательный механизм
выноса веществ из мозга в кровь. Паутинная оболочка в виде ворсинки (В)
проникает сквозь мозговую оболочку в венозный синус (С). Каналы, имеющиеся в
ворсинке, позволяют небольшим количествам ликвора проникать из подпаутинного
пространства (П) в кровь. Э — эндотелий. |
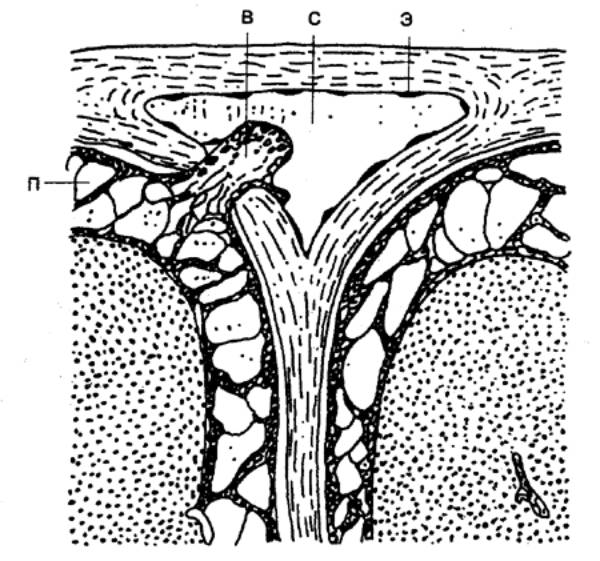
нарушить его работу) и токсические
вещества, которые все—таки в небольших количествах проникают в мозг сквозь ГЭБ.
Самый простой,
неизбирательный механизм выноса состоит в том, что избыток ликвора периодически
сбрасывается в кровь через ворсинки паутинной оболочки, выдвинутые в
просвет венозных синусов (рис. 3.78).
Ворсинки выполняют
роль клапанов, регулирующих отток: быстрое поступление ликвора в венозный синус
повышает давление в нем, ворсинка сдавливается, и отток ликвора прекращается. У
человека за 1 сут образуется примерно 500 мл ликвора, следовательно,
сопоставимое количество его оттекает ежедневно в венозную кровь.
Существуют и специфические
транспортные системы,
каждая из которых выводит из мозга против градиента концентрации конкретное
вещество или группу структурно родственных веществ:
ионы К+; мелкие
нейтральные аминокислоты; глицин, аспарагиновую и глутаминовую кислоты; пептиды
из семейства вазопрессина/окситоцина; меланостатин и энкефалины; нуклеозиды.
Эти транспортные системы необходимы для поддержания гомеостаза мозга. Так,
например, выведение глицина и K+ нормализует уровень
возбудимости ЦНС, удаление избытка меланостатина повышает активность опиоидной
системы мозга.
Создание в ходе
эволюции механизмов, выносящих в кровь пенициллины, цефалоспорины и некоторые
цитостатики, в нормальных условиях отсутствующие в мозге и применяемые при
лечении воспалительных процессов ЦНС, объясняют тем, что небольшие количества
этих веществ синтезируются микрофлорой кишечника, а также попадают в организм с
растительной пищей и проникают сквозь ГЭБ в мозг.
Важную роль в
системах, выводящих вещества из мозга в кровь, играет особый белок — гликопротеин
Р (Pgp).
Обычно он присутствует в ГЭБ в небольших количествах. При наличии в мозге
очагов воспаления или опухолей проницаемость барьера возрастает, введенные в
кровь антибиотики и цитостатики легко проникают в ЦНС. В ответ на это
происходит экспрессия гена, ответственного за синтез Pgp, его количество в
структуре ГЭБ возрастает и он быстро выводит названные вещества из мозга.
Попытки проведения
различных веществ сквозь барьер. Первый из возможных подходов состоит в том, чтобы
воздействием на ГЭБ неизбирательно и обратимо повысить его проницаемость для
любых веществ. Этого добиваются следующими нефизиологическими мерами: (а)
повышением артериального давления (в том числе локальным, в сонной артерии) на
60—90 мм рт. ст. и более, что приводит
к резкому расширению сосудов мозга; (б) повышением осмотического давления крови
путем введения в сонную артерию маннитола, арабинозы, мочевины или
гипертонического раствора NaCI, что вызывает сморщивание эндотелиоцитов; (в)
вдыханием газовой смеси, содержащей 25% СО2, что обусловливает
расширение артериол мозга и переполнение капилляров. Во всех перечисленных
случаях происходит временное нарушение плотных контактов между эндотелиоцитами
и образование водных каналов. Очевидно, что такие способы повышения
проницаемости ГЭБ не годятся для проведения лекарств в мозг человека.
Еще один подход — модификация
молекул с целью повышения их «проходимости» сквозь неизмененный барьер. Поиск
таких форм ведется по следующим направлениям: (а) повышение жирорастворимости
вещества, например, соединением его молекулы с молекулой холестерина; (б)
повышение устойчивости вещества к воздействию «энзимного барьера», например,
для пептидов это создание циклических структур или замещение L—форм аминокислот
на D—изомеры; (в) соединение препарата с веществом, для которого в ГЭБ
существует транспортная система, например, с глюкозой (создание так называемых химер);
и (г) включение изучаемого вещества в липосомы. Пока эти разработки
имеют лишь ограниченный успех.
Рассматривая механизм действия на
ЦНС любого вещества — эндогенного, присутствующего в крови, или вводимого извне
в кровь, необходимо понять, проходит ли оно сквозь ГЭБ или воздействует на мозг
иным образом, без проникновения в него.
Краткое знакомство со структурой и
функцией различных отделов центральной нервной системы позволяет сформулировать
несколько общих положений, касающихся принципов организации ее работы.
Одно из них состоит в том, что в
основе деятельности нервной системы лежит универсальный физиологический процесс
распространяющегося возбуждения — нервный импульс. Нервный импульс,
распространяясь по поверхности нервной клетки и ее отростков, выполняет в
нервной системе функцию носителя информации. Передача информации осуществляется
с помощью изменения либо последовательности нервных импульсов во времени, либо
распределения между различными клетками. Это так называемое пространственно—.
временное кодирование.
Ведущей формой нервной деятельности
являются рефлексы — закономерные реакции, возникающие в ответ на
раздражение рецепторов изменениями в окружающей или внутренней средах
организма. Рефлекторная деятельность представляет собой сложный комплекс
взаимодействующих и взаимовлияющих рефлексов. В основе рефлекторной
деятельности лежит ряд закономерностей. Это зависимость характера реакции от
типа раздражаемых рецепторов и силы действующего раздражителя, иррадиация
возбуждения, обусловленная нефункционирующими межнейронными связями ЦНС,
которые дают возможность активироваться одновременно большому числу нервных
связей.
Одна из основных закономерностей
заключается в том, что характер рефлекторной реакции способен изменяться в
зависимости от состояния тех нервных структур, через которые данная реакция
реализуется. При этом изменение возбудимости нервных структур может изменить не
только количественную, но и качественную направленность реакции. Примером тому
может служить открытое А. А. Ухтомским состояние доминанты, т. е.
появление очага возбуждения (в частности, под действием гормонов), которое приводит
к тому что стимуляция различных рецептивных полей вызывает возникновение или
усиление рефлекторной деятельности нервных структур именно этого очага.
Разные типы рефлексов осуществляются
с помощью одних и тех же мотонейронов, с использованием одних и тех же
эффекторов, т. е. мотонейроны представляют собой в этом случае общий
конечный двигательный путь. При этом правилом взаимодействия рефлексов
является у однонаправленных рефлексов взаимное усиление, у
противоположных рефлексов — взаимное торможение.
Общие закономерности тормозного
взаимодействия аналогичны закономерностям взаимного усиления. Облегчающие и
тормозные взаимные влияния между нервными центрами в процессе сочетанной
рефлекторной деятельности описаны Ч. Шеррингтоном на примере спинного мозга в
виде положительной и отрицательной одновременной индукции.
При сочетанной рефлекторной
деятельности помимо пространственного взаимодействия в нервных центрах
возникают еще и сложные явления, разворачивающиеся во времени. Еще в прошлом
столетии И. М. Сеченовым было установлено, что прекращение торможения
рефлекторной реакции сменяется так называемой отдачей — последующим
усилением по сравнению с исходной величиной. Явление отдачи рассматривают
сейчас как результат дополнительного включения более сложных центральных и
периферических путей.
Каждый рефлекторный акт всегда
является компонентом комплексной системы нервных процессов, направленных на
более эффективное выполнение определенной функции организма. Это объединение не
бывает жестко фиксированным и представляет собой динамическое взаимодействие.
Объединение относительно простых
рефлекторных механизмов и процессов в комплексы (функциональные системы),
способные уже оценивать результат рефлекторной деятельности и таким образом
регулировать эту деятельность, было изучено П. К. Анохиным. Любая часть нервной
системы является многоуровневой и построена по иерархическому принципу.
Одиночный нейрон, нейрональная цепь, нервный центр представляют собой
соподчиненные уровни системной организации, обладающей определенной
направленностью действия. Этот системный принцип встречается везде
независимо от того, идет ли речь о регуляции висцеральных функций или
организации комплексных систем корковых модулей. Системный подход наиболее
перспективен при исследовании общих аспектов поведения, таких как
взаимодействие сознания и подсознания, эмоций, научения и памяти.
Следующее положение касается
универсальности некоторых общих принципов организации нервной системы. В
качестве примера можно привести принцип доминанты, одинаково приемлемый для
спинного мозга и для колонок сенсомоторной коры. Доминанта как общий принцип
работы нервных центров определяет биологическую направленность поведения, без
учета которой глубинный анализ механизмов интегративной деятельности мозга
практически невозможен.
Сравнительно—физиологические
исследования нейронных сетей у позвоночных и беспозвоночных животных показали
принципиальное сходство их организации. Такие физиологические феномены, как суммация,
синоптическая потенциация, реципрокное торможение и ряд других имеют место
и в простых нервных системах беспозвоночных, и в ЦНС высших позвоночных.
Не меньшее число аналогий дает и
клеточный уровень организации. Достаточно напомнить, что все основные постулаты
мембранной теории разработаны на гигантских аксонах кальмара, затем повторены
на нейронах позвоночных. Сходство морфофункциональной организации нейронов и
нейронных сетей различных животных свидетельствует о возможно большем
консерватизме в области филогенетического преобразования молекулярных и клеточных
структур по сравнению с системным уровнем организации, где мы встречаемся с
явлениями параллелизма и специфичности.
Регулирующая и управляющая деятельность характеризует все
отделы нервной системы. Она распространяется на все области тела, его отделы,
органы, ткани. Относительно простыми формами такой деятельности характеризуются
структуры спинного мозга, первично управляющие соматическими и
висцеральными реакциями. Сигналы для соматических моторных реакций
(собственные, защитные, ритмические, позные и другие рефлексы мышц) формируются
спинальными мотонейронами, для висцеральных — симпатическими и
парасимпатическими преганглионарными нейронами, локализующимися в грудном и
крестцовом отделах спинного мозга. Из висцеральных реакций наиболее изучены механизмы,
обеспечивающие регуляцию уровня кровяного давления за счет изменения просвета
сосудов. По такому функциональному признаку в спинном мозгу разделяются лишь
эфферентные нейроны. Что касается чувствительных и особенно вставочных, то они
способны активизировать и соматические двигательные нейроны, и висцеральные,
управляющие сосудодвигательной функцией, регуляцией сокращений мочевого пузыря
и др.
Значительно сложнее деятельности спинного мозга рефлекторная
деятельность продолговатого мозга и мозжечка (ромбовидный мозг). Их нервные
образования (ретикулярная формация ствола, подкорковые ядра, кора мозжечка) не
получают сенсорных сигналов непосредственно от рецепторных структур и не отдают
также эфферентных сигналов непосредственно исполнительным органам. Они связаны
только с другими центрами мозга и являются, строго говоря, надстройкой над
первичной сегментарной структурой ЦНС. Осуществляемые ядрами расположенных
здесь черепных нервов рефлекторные реакции, как и в спинном мозгу, по характеру
конечного эффекта разделяются на соматические и висцеральные.
Первые направлены главным образом на поддержание позы, вторые — на обеспечение
последовательности восприятия, жевания, глотания и продвижения пищи по
пищеварительному каналу.
Ряд важнейших висцеральных функций связан непосредственно с ретикулярной
формацией продолговатого мозга. В ней имеются комплексы клеток, ритмичность
деполяризации мембран которых у одной части соответствует фазе вдоха, у другой
— фазе выдоха («вдыхательные» и «выдыхательные» нейроны). На деятельность этих
групп нейронов специфическое действие оказывает парциальное давление CO2 в крови. Чем оно выше, тем чаще
ритм, и наоборот. Эта автоматическая деятельность может изменять свою
активность под влиянием импульсов из различных рецептивных зон, а также в
определенной мере произвольным волевым усилием.
Кроме того, в продолговатом мозгу находятся и комплексы
нейронов, связанных с регуляцией сосудистого тонуса: одни из них учащают
свою деятельность при повышении артериального давления, другие — при его
понижении («сосудосуживающие» и «сосудорасширяющие» нейроны). Нисходящие пути,
образованные такими клетками, направляются в спинной мозг, где синаптически
контактируют с преганглионарными симпатическими нейронами, обладающими
сосудодвигательными функциями. Подобно дыхательным нейронам, клетки
сосудодвигательного центра обладают высокой чувствительностью к изменениям
химического состава крови.
Функции сосудодвигательного центра тесно связаны с
деятельностью группы нейронов, регулирующих работу сердца. Эти нейроны
находятся в постоянной тонической активности, которая модулируется влиянием со
стороны сосудодвигательного центра. Возбуждение клеток сердечной, области через
соответствующие ганглионарные нейроны (главным образом метасимпатической
природы) вызывает торможение деятельности сердца.
Нейронные механизмы среднего мозга обслуживают в
основном дистантные функции организма — зрение, слух, а также регулируют
определенные двигательные процессы. Нейроны последней специализации находятся
под непосредственным контролем и влиянием коры больших полушарий и подкорковых
ядер. Они образуют систему экстрапирамидных двигательных путей.
Рефлекторная деятельность промежуточного мозга,
который включает большое число различных структур, локализующихся вокруг III желудочка (главными среди них
являются таламус и гипоталамус), исключительно сложна. Все
сенсорные системы, за исключением обонятельной, проходят через ядра таламуса
перед вступлением в кору. Помимо проекционных нейронов ядер таламуса, в нем
имеются клетки, аксоны которых следуют в другие подкорковые структуры.
Комплексы таких нейронов обозначают как неспецифические ядра таламуса.
Посредством двусторонних связей с другими подкорковыми структурами таламус
регулирует взаимодействие ряда сложнорефлекторных функций, требующих интеграции
соматических и вегетативных реакций.
В этом отношении исключительно важное значение принадлежит гипоталамусу.
За счет связей со стволовыми структурами мозга и ведущей эндокринной железой — гипофизом
— гипоталамус выполняет функцию высшего центра интеграции висцеральных
функций, которые далее регулируются симпатическими, парасимпатическими и
метасимпатическими частями автономной нервной системы, а также эндокринными
влияниями. В этом случае центральные процессы, направляющие деятельность организма
на поиск удовлетворения или избегания неблагоприятных ситуаций, обозначают
понятием мотивация.
Самой новой в эволюционном отношении структурой нервной
системы является конечный мозг. Все поверхности его полушарий покрыты
корой большого мозга, образованной серым веществом. Вместе с другими
структурами нервной системы кора участвует в регуляции и координации функций
организма, играет исключительную роль в психической или высшей нервной
деятельности.
Высшая нервная деятельность представляет собой интегративную
деятельность высших отделов мозга, обеспечивающую индивидуальное поведенческое
приспособление человека и высших животных к изменяющимся условиям внутренней и
окружающей среды. При введении этого понятия И. П. Павлов определил его отличие
от низшей нервной деятельности. Последняя объединяет совокупность безусловных
рефлексов, обеспечивающих координированную деятельность организма, направленную
на поддержание постоянства внутренней среды, например уровня кровяного,
осмотического, онкотического давления.
Следовательно, высшая нервная деятельность представляет
собой аналитико—синтетическую деятельность коры и ближайших подкорковых
образований головного мозга. Она проявляется в способности выделять из
окружающей среды ее отдельные элементы и объединять их в комбинации, которые
точно соответствуют биологической значимости явлений окружающего мира.
Главная структурная особенность коры — огромное число (более 10 млрд.)
нейронов. Клетки всех слоев, которые расположены перпендикулярно по отношению к
поверхности коры и тесно связаны между собой и подкорковыми образованиями,
образуют комплекс, называемый колонкой клеток. Каждая подобная колонка
отвечает за восприятие по преимуществу одного вида чувствительности. В коре
выделяют сенсорные зоны, к клеткам которых проецируются основные
сенсорные системы (вкусовая, слуховая, зрительная и др.), а также моторные
зоны, которые посылают команды по пирамидным и экстрапирамидным путям в
ствол мозга и в спинной мозг. Клетки этих зон также посылают сигналы к
эффекторным нервным клеткам.
Одним из узловых вопросов физиологии высшей нервной
деятельности является научение. Сейчас принято различать ряд его форм:
1) облигатное, стимулзависимое (суммация, привыкание, импринтинг, подражание);
2) факультативное, эффектзависимое (условные рефлексы, классические и
инструментальные); 3) когнитивное (основанное на образной памяти, элементарной
рассудочной деятельности, вероятностном прогнозировании).
Важнейшими принципами интегративной деятельности мозга
являются принцип условного рефлекса (И. П. Павлов) и принцип
доминанты (А. А. Ухтомский),
сочетание которых обеспечивает активно—творческий и высокоадаптивный характер
целостного поведения. В качестве главного субстрата управления интегративной
деятельностью мозга сейчас рассматривают высшие ассоциативные системы; что
касается психики человека, то биологические предпосылки ее формирования зависят
от трех видов потребностей — витальных, социальных и идеальных.
Человеческий мозг характеризуется функциональной и
межполушарной асимметрией: правое полушарие работает по принципу
дедукции, т. е. сначала осуществляет синтез, а затем анализ, левое — по
принципу индукции, т. е. сначала анализирует, а затем синтезирует информацию. В
формировании речевой деятельности и мышления человека особую роль играют
активно развивающиеся в онтогенезе ассоциативные системы больших полушарий
мозга.
Барьер между кровью (гемолимфой) и ЦНС независимо возник не
менее чем в трех ветвях эволюции: у позвоночных, головоногих моллюсков и высших
членистоногих в связи с усложнением функций мозга и возросшей потребностью в
чрезвычайно строгом поддержании постоянства его внутренней среды.
У позвоночных основу гематоэнцефалического барьера
(ГЭБ) составляют эндотелиоциты капилляров мозга, соединенные между собой особого
рода контактами. Важную роль играют также вещества, секретируемые отростками
астроцитов, и энзимы, находящиеся в тканях между кровью и нейронами мозга.
Способность веществ проникать сквозь ГЭБ определяется в
первую очередь их растворимостью в жирах; для некоторых других веществ,
необходимых мозгу (например, для глюкозы и аминокислот), в барьере существуют
системы переносчиков.
На границе между кровью и мозгом есть небольшие по площади,
но очень важные участки, в которых функцию барьера выполняет не эндотелий
капилляров, а эпендима желудочков мозга. Существуют также пути «обхода»
барьера, самым известным (и, тем не менее, малоизученным) у человека является
путь, начинающийся на слизистой оболочке полости носа.
ГЭБ не только обеспечивает поддержание особого гомеостаза
мозга, но и является сложной коммуникационной системой, связывающей мозг и
периферические органы. Эти связи могут осуществляться и без проникновения
каких—либо веществ сквозь барьер (например, гормоны могут связываться с
рецепторами на внешней стороне барьера). Проникновение сквозь барьер
осуществляется в обе стороны (из крови в мозг и из мозга в кровь). Транспортные
системы, выносящие вещества из мозга, играют важную роль в поддержании его
гомеостаза. Средства, которые применяют для повышения проницаемости ГЭБ (резкое
повышение напряжения CO2 или осмотического давления плазмы
крови и т. п.), нельзя считать физиологическими — они приводят не к вскрытию,
но скорее к взлому барьера.
Айрапетянц М. Г., Вейн А. М. Неврозы в эксперименте и в клинике.
М., 1982.
Альтман Я. А. Психофизиологический анализ
поэтического вдохновения. СПб., 1994.
Анохин П. К. Биология и нейрофизиология
условного рефлекса. М., 1968.
Батуев А. С. Высшая нервная деятельность. М.,
1991.
Батуев А. С. Высшие интегративные системы мозга.
Л., 1981.
Бериташвили И. С. Избранные труды: Нейрофизиология и
нейропсихология / И. С. Бериташвили. Л., 1975.
Бернштейн Н. А. Очерки по физиологии движений и
физиологии активности. М., 1966.
Бехтерева Н. П. Здоровый и больной мозг человека.
Л., 1988.
Бехтерева Н. П. Магия мозга и лабиринты жизни.
СПб., 1999.
Бехтерева Н. П. О мозге человека: Размышления о
главном. СПб., 1994.
Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум и поведение. М., 1988.
Брёже М. Электрическая активность нервной
системы. М., 1979.
Буреш Я., Бурешова О., Хьюстон Дж. Методики и основные эксперименты по
изучению мозга и поведения. М., 1991.
Вартанян Г. А., Пирогов А. А. Эмоциональный анализатор мозга.
СПб., 1994.
Вейн А. М., Хехт К. Сон человека: Физиология и
патология. М., Берлин, 1989.
Гельгорн Э. Луфборроу Дж. Эмоции и эмоциональные
расстройства. Нейрофизиологическое исследование. М., 1966.
Громова Е.А. Эмоциональная память и ее
механизмы. М., 1980.
Дельгадо X. Мозг и сознание. М., 1971.
Дьюсбери Д. Поведение животных: Сравнительные
аспекты. М., 1981
Карамян А. И. Эволюция конечного мозга
позвоночных. Л., 1976.
Кассиль В. Г. Пищевое поведение в онтогенезе. Л.,
1990.
Кассиль Г. Н. Наука о боли. М., 1975.
Ковбаса С. И., Ноздрачев А. Д. Информационные характеристики
систем нейронов. Л., 1990.
Ковбаса С. И., Ноздрачев А. Д.,
Ягодин С. В. Анализ
взаимосвязи нейронов. Л., 1984.
Костюк П. Г. Физиология центральной нервной
системы. Киев, 1977.
Котляр Б. И., Шульговский В. В. Физиология центральной нервной
системы. М., 1979.
Кругликов Р. И. Нейрохимические механизмы обучения
и памяти. М., 1981.
Крушинский Л. В. Биологические основы рассудочной
деятельности. М., 1986.
Куффлер С., Николе Дж. От нейрона к мозгу. М., 1979.
Кэндел Э. Клеточные основы поведения. М.,
1980.
Лапицкий В. П. Головные ганглии и двигательная
активность насекомых. Л., 1990.
Мак—Фарленд Д. Поведение животных: Психобиология,
этология и эволюция. М., 1988.
Милейковский Б. Ю., Ноздрачев А. Д. Торможение двигательной активности.
Стволовые механизмы. СПб., 1998.
Ноздрачев А. Д., Поляков Е. Л. Анатомия кошки. СПб., 1998.
Ноздрачев А. Д., Поляков Е. Л. Анатомия лягушки. М., 1994.
Ноздрачев А. Д., Поляков Е. Л„
Гнетов А. В.
Исследование функций головного мозга: некоторые современные методы. Л., 1987.
Ноздрачев А. Д., Поляков Е. Л.,
Лапицкий В. П., Осипов Б. С., Фомичев Н. И. Анатомия беспозвоночных: пиявка, прудовик,
дрозофила, таракан, рак.
Ноздрачев А. Д., Чумасов Е. Н. Периферическая нервная система.
СПб., 1999.
Ноздрачев А. Д.. Чернышева М. П. Висцеральные рефлексы. Л., 1989.
Поляков Г. И. Проблема происхождения рефлекторных
механизмов мозга. М., 1964.
Раус С. Устройство памяти. От молекул к
сознанию. М., 1995.
Серков Ф. Н. Корковое торможение. Киев, 1986.
Симонов П. В. Мотивированный мозг: высшая нервная
деятельность и естественнонаучные основы общей психологии. М.,1987.
Симонов П. В. Эмоциональный мозг. М., 1981.
Слоним А. Д. Инстинкт: загадки врожденного
поведения организмов. Л., 1967.
Соколов Е. Н. Нейронные механизмы памяти и обучения.
М., 1981.
Сравнительная физиология животных. В
3 т. / Под ред. Л. Проссера. М., 1977.
Ухтомский А. А. Доминанта. М.—Л.,
1966.
Физиология поведения.
Нейробиологические закономерности / В. П. Бабминдра и др. Л., 1987.
Физиология поведения. Нейрофизиологические
закономерности / А. С. Батуев, А. Д. Солоним, П. В. Симонов и др. Л., 1986.
Функциональные системы организма / Под ред. К. В. Судакова.
М., 1987.
Хайнд Р. Поведение животных. Синтез этологии
и сравнительной психологии. М., 1975.
Хьюбел Д. Глаз, мозг, зрение. М., 1990.
Черниговский В. Н. Интероцепция: Сборник. Л., 1985.
Шаде Д., Форд Д. Основы неврологии. М., 1976.
Шаповалов А: И. Механизмы синаптической передачи.
Избранные труды. СПб., 1997.
Шеперд Г. Нейробиология. В 2 т. М., 1987.
Эволюционная физиология. В 2—х ч. /
Отв. ред. Е. М. Крепе. Л., 1983. Ч. 2. Руководство по физиологии.