2 РЕГУЛИРОВАНИЕ ЖИЗНЕННЫХ ФУНКЦИЙ ОРГАНИЗМА
ВЕХИ ИСТОРИИ
1637 — Р. Декарт (R. Descartes, Франция) сделал попытку проникнуть
в сущность произвольных и непроизвольных движений. Дал первое описание рефлекса
(но не использовал этого слова).
1784 — И. Прохазка
(J. Prochaska, Чехия) развил представление о
рефлексе (и предложил этот термин), рефлекторной дуге, нервной системе как
посреднике между внешней средой и организмом.
1832 — М. Голл и
И. Мюллер (М. Goll, J. Muller, Германия) изложили учение о рефлексе.
1855 — К. Бернар
(С. Bernard, Франция) предположил, что железы выделяют в кровь
«внутренние секреты», оказывающие действие на весь организм. В 1857 г. открыл существование
постоянства внутренней среды организмов и ее значение для поддержания жизни.
1863 — И. М.
Сеченов (Россия) в книге «Рефлексы головного мозга» распространил принцип
рефлекторной реакции на психическую деятельность и поведение человека.
1897 — Ч.
Шеррингтон (Ch. Sherrington, Великобритания) ввел понятие «синапс».
1902 — в докладе
из лаборатории И. П. Павлова впервые заявлено о существовании условных
рефлексов.
1902 — У. Бейлисс
и Э. Старлинг (W. Bayliss, E. Starling, Великобритания) открыли секретин —
первый из гормонов, и таким образом, основали эндокринологию. В 1905 г. по
предложению У. Харди (W.Hardy, Великобритания) Старлинг назвал новую группу веществ
«гормонами».
1906 — Ч.
Шеррингтон разработал концепцию интегративных процессов в нервной системе.
1908 — И. И.
Мечников (Россия) и П. Эрлих (Р. Ehrlich, Германия) — Нобелевская премия за
создание теории иммунитета.
1923 — А. А.
Ухтомский (СССР) создал учение о доминанте.
1928 — Э.
Шаррер (E. Scharrer, Германия) ввел понятие «нейросекреторная клетка», но его
взгляды получили признание только в 1960—е годы.
1929 — У. Кеннон (W. Cannon, США) ввел понятие о гомеостазе.
1930—е — П. К.
Анохин (СССР) высказал идею об «акцепторе результата действия» и, таким
образом, открыл существование обратной связи в физиологических процессах (но не
использовал этого термина).
1931 — У. фон Эйлер и Дж. Гаддам (U. v. Euler, J. Gaddam, Швеция) открыли субстанцию Р — первый
из регуляторных пептидов.
1932 — Ч.
Шеррингтон и Э. Эдриан (E. Adrian, Великобритания) — Нобелевская премия за открытие
роли рецепторов, чувствительных и двигательных нервов, передачи информации в
виде электрических импульсов.
1936 — О. Лёви (О. Loewi, Австрия) и Г. Дейл (Н. Dale, Великобритания) — Нобелевская
премия за открытие механизма синаптической передачи.
1936 — Г. Селье
(Н. Selye, Канада) открыл феномен стресса и описал общий адаптационный синдром и
болезни адаптации.
1941.— Н. А. Бернштейн (СССР) в трудах по
физиологии движений предвосхитил некоторые последующие идеи кибернетики, создал
схему рефлекторного кольца.
1947 — Б.
Усай (В. Houssay, Аргентина) — Нобелевская премия за исследования гормонов
передней доли гипофиза.
1948 — Н. Винер (N. Wiener, США) создал новую науку —
кибернетику.
1949 — В.
Хесс (W. Hess, Швейцария) — Нобелевская премия за открытие гипоталамических центров
регуляции автономных функций.
1949 — в книге У. Кеннона и А. Розенблюта (A. Rosenblueth, Мексика) описан закон денервации,
повышение чувствительности денервированных структур.
1950 — Э.
Кендалл (Е. Kendall, США), Т. Рейхштейн (Т. Reichstein, Швейцария) и Ф. Хэнч (Ph. Hench, США) — Нобелевская премия за
открытие кортикостероидов.
1955 — В. Дю Виньи (V. Du Vigneaud, США) — Нобелевская премия по химии
за впервые осуществленный синтез пептидного гормона — окситоцина.
1971 — Э. Сазерленд (Е. Sutherland, США) — Нобелевская премия за
открытие вторичных мессенджеров (посредников).
1977 — Р. Гиймен и Э. Шалли (R. Guillemin, A. Schally, США) — Нобелевская премия за
открытие рилизинг—гормонов гипоталамуса.
1978 — А. Пирс (A. Pearse, Великобритания) открыл
существование диффузной эндокринной системы (APUD—системы).
1979 — А.
Кормак (A. Cormack, США) и Г. Хаунсфилд (G. Hounsfield, Великобритания) — Нобелевская
премия за разработку метода компьютерной томографии.
1980 — А. Д. Ноздрачев (СССР) ввел понятие
метасимпатическая нервная система (МНС) и определил ее роль в регуляции
висцеральных функций.
1986 — С. Коэн (S. Cohen, США) и Р. Леви—Монтальчйни (R. Levi—Montalcini, Италия—США) — Нобелевская премия
за открытие пептидных факторов роста.
Выживание организма в условиях непрерывно меняющейся среды
обусловлено способностью живых систем сохранять свое стационарное состояние.
Для обеспечения такого состояния у всех организмов — от простых до самых
сложных — существуют различные анатомические, физиологические и поведенческие
приспособления. Все они направлены на поддержание постоянства внутренней среды.
Положение же о том, что именно постоянство внутренней среды определяет
оптимальные условия для жизни и размножения организма, было высказано великим
французским физиологом Клодом Бернаром в 1857 г.
Организм находится в состоянии непрерывного приспособления к
условиям окружающей среды. Его существование было бы невозможно, если бы он не
реагировал адекватным образом на недостаток кислорода или пищи, избыток
углекислого газа или других многочисленных вредоносных агентов. Каждый животный
организм представляет собой сложную обособленную систему, внутренние силы
которой каждый момент, покуда она существует как таковая, уравновешиваются с
внешними силами окружающей среды... Таким образом, вся жизнь от простейших до
сложнейших организмов... есть длинный ряд все усложняющихся до высочайшей
степени уравновешиваний внешней среды» (И. П. Павлов). Совокупность
физиологических процессов, обеспечивающих это уравновешивание организма со
средой, относится к явлениям регуляции. В основе явлений регуляции лежит
взаимосвязь всех органов и функциональных систем организма между собой.
Для обеспечения относительно стабильной активности организма
необходима регуляция систем его жизнеобеспечения на
|
|
|
|
Рис.
2.1 Система управления с обратной
связью в биологических объектах — положительной (А) и отрицательной (Б) Стрелками показаны направления
воздействий. |
|
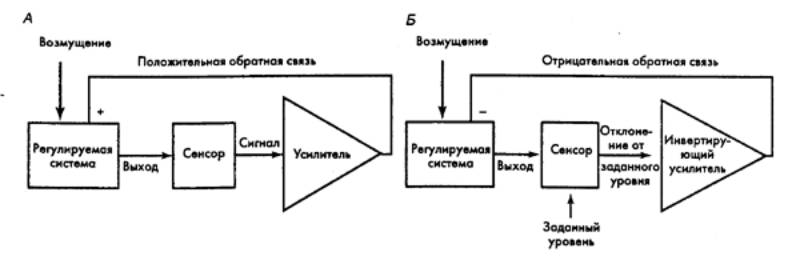
всех уровнях — от молекулярного до организации и условиям жизни данного
вида. В этом отношении млекопитающие лучше оснащены, нежели беспозвоночные или
простейшие.
Существующие у живых организмов механизмы и способы
регуляции функций имеют ряд принципиально общих черт с подобными устройствами в
неживых системах, таких как машины. И у тех, и у других стабильность
достигается благодаря существованию определенных форм управления. Эти формы
управления Норберт Винер назвал в 1948 г. кибернетикой (от греч. kybernetike —искусство управления).
Именно кибернетика занимается сейчас изучением общих закономерностей
регулирования в живых и неживых системах.
Для объяснения механизмов, лежащих в основе функционирования
биологических регуляторных систем, как правило, прибегают к привлечению
математических моделей теории управления. Следовательно, прежде чем
рассматривать процесс регулирования жизненных функций, необходимо в сугубо
общих чертах ознакомиться с некоторыми положениями теории управления.
Как известно, живые системы рассматриваются как открытые
системы. Для поддержания своего состояния они нуждаются в непрерывном
обмене веществами с окружающим миром и находятся в динамическом равновесии с
ним. Для этого необходим постоянный приток энергии. Равновесие устанавливается
только после гибели организма, когда он становится термодинамически стабильным
по отношению к среде.
В любой управляющей системе мерой эффективности
является степень отклонения регулируемого параметра от оптимального (должного)
уровня и скорость возвращения к этому уровню. Еще одним непременным условием
работы любого механизма является свобода колебаний. Эти условия исключительно
важны, так как именно колебания активируют систему управления и возвращают
переменную к оптимальной величине. Такие системы, предусматривающие соединение
компонентов, при котором выход регулируется входом, действуют по принципу
обратной связи.
Принято рассматривать два вида обратной связи —
отрицательную и положительную. В открытых живых системах более
распространена первая. Принцип ее организации легко рассмотреть на примере
поддержания постоянства температуры тела, регуляции артериального кровяного
давления, регуляции напряжения кислорода, уровня кортикостероидов или других
гормонов в крови. Остановимся на регуляции кортикостероидов надпочечником. В
этом случае модулятор состоит из трех компонентов — детектора (гипоталамуса),
регулятора (гипофиза) и эффектора (коры надпочечника). При нарушении равновесия
системы возникает ряд последствий, которые приводят к устранению этого нарушения
и к возвращению системы в исходное состояние (рис. 2.1).
В отличие от отрицательной, положительная обратная связь у
биологических объектов встречается относительно редко. Связь эта приводит к
нестабильности системы и экстремальным ситуациям, сопровождаясь такими
последствиями, которые углубляют исходное состояние. Наличие положительной
обратной связи легко рассмотреть на примере распространения нервного импульса
по волокну. Как известно, деполяризация нейрона повышает проницаемость мембраны
для Na+ Ионы Na+ входят в аксон через мембрану и углубляют деполяризацию. Деполяризация
в свою очередь приводит к возникновению потенциала действия. Положительная
обратная связь в этом случае играет роль усилителя ответа, величину которого
ограничивают иные механизмы.
Рассмотренные механизмы обратной связи являются простейшими;
реально в организме используются значительно более сложные регуляторные
устройства, включающие дополнительные детекторы типа физиологических систем
раннего предупреждения, или в случае отказа исполнительных структур включаются
дополнительные эффекторы. Действуют эти системы связи на разных уровнях
управления. Множественные детекторы и эффекторы обеспечивают дополнительную
надежность регуляции таких исключительно важных показателей, как, например, артериальное
давление.
2.1. СОВЕРШЕНСТВОВАНИЕ РЕГУЛЯТОРНЫХ
МЕХАНИЗМОВ В ПРОЦЕССЕ ЭВОЛЮЦИИ
Древнейшая форма взаимодействия между клетками
многоклеточных органов — химическое влияние, осуществляющееся
посредством продуктов обмена веществ, которые выделяются в жидкости организма.
В этом случае все многообразие физиологически активных соединений, образующихся
в процессе жизнедеятельности органов, тканей и клеток, включая продукты распада
белков, углекислый газ, электролиты и др., лежит в основе гуморальных механизмов
связи между органами.
Межклеточное (креаторное) взаимодействие через
микроокружение тканей — один из наиболее эволюционно старых способов регуляции
в организме. Оно осуществляется макромолекулами, несущими информацию,
необходимую для регулирования внутриклеточного синтеза определенных молекул
белка для обеспечения дифференцировки, роста, развития и объединения клеток в
ткани. К веществам, обеспечивающим такого рода местную гуморальную регуляцию,
можно отнести кейлоны — белки или гликопротеиды, способствующие
подавлению деления клеток и синтеза ДНК. Возможно, нарушение креаторных связей
обусловливает патогенез опухолевого роста и других заболеваний.
Местные регуляторные эффекты метаболитов неспецифичны и
сводятся к регуляции обменных процессов и функции органов — источников их
возникновения. Так, избыток молочной и пировиноградной кислоты при интенсивной
мышечной работе угнетает сократительную способность мышц и в то же время
приводит к увеличению в них кровотока (рабочей гиперемии), интенсификации метаболизма
этих недоокисленных продуктов.
Гуморальные связи, общие и для мира животных, и для
растений, характеризуются рядом особенностей. Во—первых, относительно медленным
распространением химического вещества. Во—вторых, отсутствием точного адреса,
по которому следует соединение, поступающее в кровь, лимфу или тканевую
жидкость. Его действие не локализовано, не ограничено определенным местом и,
следовательно, диффузно. И наконец, в—третьих, малой надежностью связи,
поскольку вещество высвобождается в малых количествах и обычно быстро
разрушается или выводится из организма.
Кроме того, физиологически активные
вещества, разносясь током крови по всему, организму, только в определенных
местах, в результирующих органах или клетках—мишенях, вызывают целенаправленные
специфические реакции при взаимодействии с эффекторами или соответствующими
рецепторными образованиями.
В процессе эволюции животного мира
механизмы гуморальной регуляции постепенно дополнялись более сложными и более
совершенными механизмами нервной регуляции функций. Более того, по мере
совершенствования живых систем гуморальные связи все более подчиняются нервному
влиянию. Устанавливается фактически неразделимое взаимодействие, между
гуморальными и нервными компонентами регуляторного процесса — единая система
взаимосвязанных нейрогуморальных отношений.
Доминирование у высокоорганизованных
существ нервных механизмов регуляции функций не означает редукции гуморальной
системы связи. На следующем этапе развития живых существ появляются специальные
органы — эндокринные железы, вырабатывающие гуморально действующие
вещества — гормоны. Их действие зачастую весьма специализировано.
Доказано также, что в окончаниях
нервных волокон, образующих синаптические контакты с мышцами или другими
нервными клетками, высвобождаются посредники, переносчики нервного влияния, или
медиаторы (трансмиттеры), которые действуют на специализированные
системы рецепторов и тем самым осуществляют исполнительный акт. Влияние
гуморальных веществ на клетки осуществляется и непосредственно на рецепторы,
регулирующие проницаемость ионных каналов поверхностных мембран, и косвенно
через ряд промежуточных инстанций, и, в частности, через образование
циклических нуклеотидов (аденозин— или гуанидин—5 монофосфат) — внутриклеточных
посредников.
Известны четыре системы вторичных
посредников: 1 — аденилатциклаза — циклический аденозинмонофосфат (цАМФ), 2
— гуанилатциклаза — циклический гуанозинмонофосфат (цГМФ), 3 — фосфолипаза С —
инозитолтрифосфат (ИФ), 4 — ионизированный кальций.
Нервная регуляция как высший этап
развития приспособления организма к меняющимся условиям среды. В прошлом некоторые исследователи
противопоставляли нервную регуляцию функций гуморальной. Сторонники
гуморального направления игнорировали и сводили к минимуму роль нервных связей,
в то время как представители нервной теории стремились опровергнуть наличие
сколько—нибудь существенного химического взаимодействия между тканями
организма. Современная физиология полностью отвергает противопоставление
отдельных видов регуляции.
В многочисленных экспериментах
показано, что разнообразные продукты обменных реакций могут действовать как
раздражители не только непосредственно на мембраны клеток, но и на нервные
окончания — хеморецепторы, вызывая рефлекторным путем определенные
физиологические и биохимические сдвиги.
Так, многие передатчики нервного
влияния — медиаторы, выполнив свою основную роль и избежав ферментативной
инактивации или обратного захвата нервными окончаниями, попадают в кровь,
осуществляя дистантное (немедиаторное) действие. Проникая через
гистогематические барьеры, они поступают в органы и ткани и регулируют их
жизнедеятельность. Состояние самой нервной системы зависит не только от
информации из окружающей и внутренней среды, но и от снабжения кровью и от
различных ингредиентов внутренней среды.
При этом имеет место теснейшая
взаимосвязь и взаимообусловленность нервных и гуморальных процессов. Так,
нейросекреторные клетки гипоталамических ядер являются местом трансформации
нервных стимулов в гуморальные, а гуморальных в нервные. Помимо разнообразных
медиаторов в мозгу синтезируются многочисленные пептиды и другие активные
соединения, принимающие участие в регуляции деятельности головного и спинного
мозга, а при поступлении в кровь — всего организма. Таким образом, и мозг
тоже можно назвать эндокринной железой.
Физиологическая активность жидких
сред организма обусловлена в значительной степени соотношением электролитов и
микроэлементов, состоянием синтезирующих и расщепляющих ферментных систем,
наличием активаторов и ингибиторов, образованием и распадом сложных белково—полисахаридных
комплексов, связыванием и высвобождением субстратов несвязанных форм и т. д.
Важную роль в нейрогуморальной
регуляции функций играют гормоны, разнообразные продукты промежуточного обмена,
объединяемые под названием метаболиты, тканевые гормоны,
гипоталамические нейрогормоны. Они меняют чувствительность клеток (количество и
сродство рецепторов) к гуморальным и нервным влияниям, модулируют регуляторные
сигналы за счет сдвигов проницаемости мембран, их электрических зарядов,
рецепции, образования вторичных посредников, интенсивности энергетического
метаболизма и кровоснабжения тканей. Образуясь в неспецифических клетках,
большинство биологически активных веществ способно проникать из межклеточной
среды в кровь и, таким образом, наряду с основным местным регуляторным влиянием
оказывать региональные и даже генерализованные эффекты.
Какие же преимущества обеспечивает
способ регуляции функций, осуществляемый с преимущественным участием нервного
аппарата? В отличие от гуморальной связи нервная связь, во—первых, имеет точную
направленность к определенному органу и даже группе клеток и, во—вторых, через
нервные проводники связь осуществляется с гораздо большей скоростью, в сотни
раз превышающей скорость распространения физиологически активных веществ.
Наряду с кабельным способом управления по принципу «абонент—ответ», как на
телефонной станции, центральный аппарат нервной системы с преобладающими
интегративными промежуточными нейронами обеспечивает вероятностный принцип
управления, гибко приспособленный к непрерывно меняющейся обстановке и
обеспечивающий детермированные исполнительные реакции.
2.2. ХАРАКТЕРИСТИКА ГУМОРАЛЬНЫХ
МЕХАНИЗМОВ РЕГУЛЯЦИИ
В процессе эволюции при образовании многоклеточных
организмов между отдельными клетками возникли функциональные связи; с
усложнением их строения становились все более необходимыми механизмы,
обеспечивающие взаимодействие между клетками.
Для осуществления коммуникации и координации между клетками,
для поддержания стабильности внутренней среды организма — гомеостаза — и
осуществления основных жизненных функций в эволюции возникли две основные
системы — эндокринная и нервная, работающие во взаимодействии
между собой при осуществлении интегративной деятельности.
Учение об эндокринной регуляции создано во второй половине XIX в. А. Бертольдом, Ш. Броун—Секаром и др. Эндокринная система
осуществляет свое влияние" гуморально посредством ряда веществ с
регуляторным действием — гормонов. Первый из гормонов — секретин был открыт У.
Бейлиссом и Э. Старлингом в 1902 г. В
1905 г. У. Харди
предложил термин «гормон»(от греч. hormao — приводить в движение, побуждать).
Гормоны обнаружены у растений и животных разных уровней
развития, однако наибольшей сложности и многообразия система гормональной
регуляции достигает у позвоночных животных.
Гормоны служат посредниками, которые вместе с нервной
системой в составе единого интегративного регуляторного аппарата координируют
самые разнообразные функции организма — метаболизм, размножение, иммунитет,
сезонные циклы активности и т. д.
По химической природе гормоны разнообразны и относятся к
двум основным группам: первая представлена стероидами, во вторую входят производные
аминокислот, пептиды, белки.
Гормоны вырабатываются в специализированных органах — эндокринных
железах, не имеющих выводных протоков (в отличие от зкзокринных желез), или
в группах клеток с эндокринной функцией в пределах разных органов (например, в
островковых клетках поджелудочной железы, в специализированных клетках гонад,
вырабатывающих половые гормоны, и т. д.), а также в эндокринных клетках,
встречающихся в слизистой оболочке кишки, в сердце и других органах. Большая
часть эндокринных желез имеет эпителиальное происхождение.
Гормоны поступают во внутреннюю среду организма — кровь,
лимфу и другие тканевые жидкости — и разносятся по всему организму.
Регулирующий эффект они оказывают путем воздействия на рецепторы эффекторных
органов— или тканей—мишеней, которое передается на сложные клеточные структуры
— мембраны, ферментные системы (см. также разд. 2.2.4). На эти органы гормоны
оказывают специфическое действие, которое не могут проявить другие вещества.
При этом эффект вызывается во многих случаях при наличии гормона в чрезвычайно
низких концентрациях — достаточно нескольких нано—, а иногда даже
пикограммов вещества. В то же время клетки других тканей не реагируют на
наличие гормона в циркулирующих жидкостях.
Таким образом, характерными особенностями гормонов являются
их выработка в специализированных железистых клетках, выведение в циркулирующие
жидкости организма, дистантное действие и способность к вызыванию специфической
активности в низких концентрациях.
Существует и ряд других регуляторных веществ, которые иногда
также относят к гормонам. Это так называемые гормоны местного действия,
или тканевые гормоны, вырабатываемые паракринными клетками и
обеспечивающие быструю регуляцию тканевых процессов. Эти вещества могут
достигать своих органов—мишеней диффузией через тканевую жидкость, без переноса
кровью. К этой группе относятся простагландины — соединения широкого спектра
физиологического действия, вазоактивные кинины, некоторые биогенные амины,
например, серотонин, гистамин и т. д.
Ведущую роль в координации и интеграции жизнедеятельности
клеток, тканей, органов и организма в целом имеет централизованная система
управления основными взаимосвязями, компонентами которой являются нервная и
эндокринная системы.
2.2.1. Основные особенности эволюции
гормональных регуляторных механизмов
В процессе эволюции, задолго до появления специализированных
эндокринных желез, вырабатывающих гормоны, появилась способность нервных клеток
продуцировать секрет, содержащий гормоны, которые поступают в гуморальную среду
организма и оказывают регулирующее действие на ряд функций. Это явление было
названо нейросекрецией. Гормоны, вырабатываемые нервными клетками, часто
называют регуляторными пептидами, или нейрогормонами.
Медиаторы также являются продуктами нервных клеток и отличаются от
нейрогормонов тем, что выделяются окончаниями аксонов в синаптическую щель. В
настоящее время показано, что одни и те же вещества могут функционировать и в
качестве медиатора, и в качестве гормона. Например, катехоламины —
адреналин и норадреналин, вырабатываемые в мозговом веществе надпочечников,
обычно именуются гормонами в связи с их функцией. Однако эти же вещества
обнаруживаются в симпатических нервных окончаниях, и в этом случае они играют
роль медиаторов (см. также разд. 5.2.2).
У беспозвоночных животных преобладающее множество гормонов
вырабатывается клетками нервных ганглиев. Так, уже у кишечнополостных имеется
примитивная нервная система; у гидры нервные клетки функционируют как
нейросекреторные, и их гормоны регулируют развитие и рост.
У более высокоорганизованных беспозвоночных животных — у
первичноротых (черви, моллюски) и вторичноротых (иглокожие, оболочники) —
большое значение в регуляторных механизмах имеют нейросекреторные элементы и их
гормоны, однако появляются также железы эпителиального происхождения. Весьма
сложна система гормональной регуляции у насекомых, которая кроме
нейросекреторных центров включает эндокринные железы первого порядка,
регулируемые нейрогормонами, и эндокринные железы второго порядка, функция которых
определяется гормонами эндокринных желез первого порядка; в свою очередь, их
гормоны регулируют функцию органов и тканей—мишеней.
Нейросекреторные центры имеются в мозгу и у всех позвоночных животных;
гормоны, вырабатываемые нейросекреторными клетками, оказывают широкое
регулирующее влияние, и этот путь регуляции является наиболее древним.
В промежуточном мозгу, в гипоталамусе, всех
позвоночных животных установлено наличие нейросекреторных клеток, гормоны
которых либо оказывают специфическое регулирующее влияние непосредственно на
органы— или ткани—мишени (антидиуретический гормон — АДГ, окситоцин и др.,
древний путь регуляции), либо воздействуют на эндокринные железы, в частности
на гипофиз (гипоталамические рилизинг—гормоны), стимулируя синтез
и секрецию соответствующих гормонов. Кроме рилизинг—гормонов (либерины)
в гипоталамусе вырабатываются ингибирующие факторы (статины), которые
совместно с либеринами регулируют функцию железистых клеток гипофиза. Эта
система получила название гипоталамо—гипофизарной; в этом случае
наиболее четко выражены органическая взаимосвязь и взаимодействие нервной и
эндокринной систем.
Гормоны железистой части гипофиза, называемой аденогипофизом, оказывают
влияние на эндокринные железы второго порядка, вырабатывающие свои гормоны,
которые регулируют различные функции организма. Часть гормонов гипофиза
оказывает непосредственное влияние на органы— и ткани—мишени. Кроме того, ряд
эндокринных желез функционирует без регулирующего влияния гормонов гипофиза.
Среди регуляторных гипоталамических
пептидов кроме упомянутых выше гормонов открыты эндорфины и энкефалины.
Эти вещества, выполняют и медиаторную функцию, оказывают модулирующее влияние
на ряд медиаторных систем. Некоторые из регуляторных гипоталамических пептидов
обнаружены не только в нейронах головного мозга, но также в кишечнике,
например соматостатин, субстанция Р, вазоактивный интестинальный
полипептид (ВИП) и др. Они действуют как гормоны желудочно—кишечного тракта.
Некоторые из этих пептидов, первоначально обнаруженные в желудочно—кишечном
тракте, найдены в коре головного мозга, например, холецистокинин, влияющий на
выделение жёлчи и секрецию панкреатического сока. Холецистокинин обнаружен в
головном мозгу, где он участвует в регуляции пищевого поведения.
Таким образом, для ряда гормонов в
настоящее время установлена множественная локализация биосинтеза.
Согласно гипотезе Пирса
(1978), клетки, вырабатывающие эти пептиды, образуют так называемую диффузную
эндокринную систему (APUD — англ.:Amine Precursors Uptake and Decarboxylation). Особенностями этих клеток
являются высокое содержание аминов, способность к захвату их предшественников и
последующему декарбоксилированию, а также способность вырабатывать биогенные
амины и пептиды. Однако не все положения гипотезы Пирса находят подтверждение;
в частности, не доказано происхождение всех элементов диффузной эндокринной
системы из одних и тех же зачатков, как это предполагал автор.
При попытке объяснить обнаружение
ряда веществ, определяемых у высших животных как гормоны, в различных системах
организма, а также нахождение тех же или чрезвычайно близких по химической
структуре веществ у многих беспозвоночных и даже у растений была высказана
гипотеза о весьма раннем появлении в эволюции молекул, способных выполнять
широкие регуляторные функции. Возможно, что первоначально возникли
ферментные молекулы, которые на более поздних этапах эволюции превратились в
гормоны. Так, например, стероидные гормоны обнаружены у растений и животных,
причем животные синтезируют стероиды из холестерина. Однако не все стероиды,
которые у высокоорганизованных животных стали выполнять функции гормонов,
существовали на низших ступенях эволюции; часть из них создавалась по мере
возникновения новых жизненных запросов в ходе дальнейшей эволюции из
предшественников.
Многие пептидные гормоны
также широко распространены и произошли от других белков, которые существовали
на тех стадиях эволюции, когда еще не существовало эндокринной системы.
Например, инсулин, продуцируемый бета—клетками поджелудочной железы у
позвоночных, обнаруживается у них также в кишке и мозгу. Инсулин или
инсулиноподобное вещество обнаружены в нервной системе ряда
высокоорганизованных беспозвоночных (моллюсков, насекомых), а также у
простейших (инфузорий) и даже у бактерий. По—видимому, предшественники
полипептидных гормонов, из которых в процессе дальнейшей эволюции выработались
современные гормоны, уже существовали у животных самых ранних ступеней
эволюции. Многие вещества, которые в настоящее время относят к гормонам,
возникли и у растений.
Возникновение эндокринных желез и
эндокринной регуляторной системы, происшедшее независимо у беспозвоночных и
позвоночных животных, явилось ароморфозом (по А. Н. Северцову) —
эволюционным преобразованием строения и функций организмов, ведущим к
морфофизиологическому прогрессу. Это дало преимущество данным группам в
поддержании гомеостаза, повышении их адаптивных возможностей и имело значение
для прогрессивной эволюции этих форм. Очевидно, здесь были использованы
молекулы, которые существовали и имели широкие регуляторные функции, являясь
ферментами, медиаторами и т. д. В эндокринной системе эти вещества стали
выполнять функции гормонов.
На ранних этапах эволюции позвоночных многие гормоны
обладают весьма широким и разнообразным спектром действия; у более высокоорганизованных
позвоночных для ряда гормонов этот спектр более узок и специфичен (например,
пролактин, кортикостероиды и др.; см. разд. 6.2). Постепенно происходит также
все большая концентрация структур центральных и периферических отделов
эндокринной системы. В пределах отдельных эндокринных желез наблюдается не
только концентрация однородных элементов, но и объединение в одном органе
различных железистых элементов, вырабатывающих разные гормоны. Такое
объединение оказывается полезным для более эффективной работы железы.
2.2.2. Регуляция функций эндокринной
системы
Гормоны функционируют в качестве элементов регулирующих
цепей, поэтому существенно, чтобы они не накапливались в организме и их уровень
строго регулировался. Накоплению гормонов препятствуют их инактивация в
эффекторных органах (главным образом в печени) и последующее выведение с мочой.
Действие многих гормонов может блокироваться благодаря секреции гормонов,
обладающих антагонистическим эффектом. Процессы синтеза и секреции гормонов
регулируются с помощью обратной связи, и этот вид управления может
осуществляться на разных уровнях одновременно.
Для обеспечения гомеостаза и поддержания на оптимальном
уровне всех параметров внутренней среды организма большое значение имеет
включение в систему регуляции различных метаболитов (жирные кислоты,
глюкоза, аминокислоты, ионы и т. д.). Так, например, ионы Са2+,
поступающие в железы, выполняют роль регуляторов секреторного процесса; глюкоза
стимулирует гликолиз и ингибирует образование неэстерифицированных жирных
кислот. От ее уровня зависит секреция инсулина и т. д. Гормоны и продукты
метаболизма могут подавлять выделение какого—либо гормона, действуя по принципу
отрицательной обратной связи. Например, инсулин снижает концентрацию сахара в
крови, а глюкагон, действуя как антагонист инсулина, повышает ее. Конечный
результат определяется соотношением этих двух эффектов.
Существует также регуляция и после выделения гормона в
кровь. Период полураспада (время, необходимое для расщепления половины
имеющегося гормона) для многих гормонов составляет лишь несколько минут.
Как уже указывалось, функционирование эндокринной системы
осуществляется в тесном взаимодействии и взаимовлиянии с нервной системой. Это
положение легко прослеживается на примере гипоталамуса. Гипоталамус
получает информацию из внешней и внутренней среды. Из внешней среды по
сенсорным системам сигналы поступают в кору больших полушарий и другие отделы
мозга. В переработанном виде они передаются в гипоталамус, который интегрирует
всю информацию, получаемую из высших отделов мозга и непосредственно из
внутренней среды. В результате этих сигналов в гипоталамусе вырабатываются
регуляторные гормоны, включающиеся в общую систему эндокринной регуляции. Кроме
того, гипоталамус контролирует деятельность симпатической и парасимпатической
нервной системы и с их помощью регулирует деятельность периферических
эндокринных желез и органов—мишеней.
2.2.3. Функциональное значение
гормонов
Гормоны оказывают широкое регулирующее влияние на различные
функции организма. Выделяют три основные функции гормонов — они
обеспечивают: развитие организма; адаптацию физиологических систем (т. е.
способность органов и тканей изменять активность в зависимости от потребности в
ней); поддержание важнейших физиологических показателей на постоянном уровне
(гомеостатическая функция).
Различные биохимические реакции могут протекать правильным
образом лишь в присутствии одного или нескольких гормонов, хотя при увеличении
их концентрации реакция не ускоряется. В этих случаях проявляется пермиссивное
(разрешающее) действие гормона, т. е. сам гормон не влияет на данную систему,
но обеспечивает возможность нормального действия другого гормона. Например,
тироксин обладает пермиссивным действием по отношению к гормонам, регулирующим
рост.
Еще одной важной особенностью действия некоторых гормонов
является синергизм, т. е. усиление действия одного гормона под влиянием
другого.
|
|
|
Рис. 2.2 Гормональные взаимодействия и механизм обратной
связи в эндокринной системе. Сплошными стрелками обозначены стимулирующие
воздействия, пунктирными — ингибирующие. |
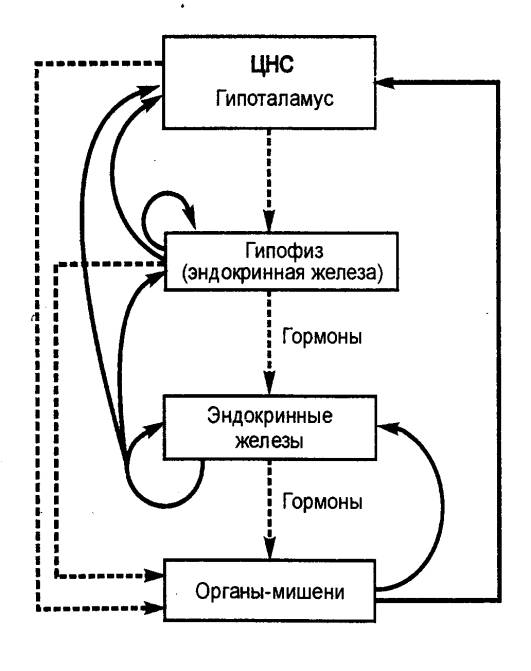
Гормоны являются элементами регуляторных систем (рис. 2.2).
С этой точки зрения они разделяются на две группы. В одной, которая включает
адреналин, норадреналин, альдостерон, АДГ и другие гормоны, скорость их
секреции и концентрация в плазме претерпевают значительные колебания; при этом
скорость секреции приспосабливается к меняющейся ситуации. Концентрации
гормонов другой группы, например тироксина, в норме колеблются мало.
Гормон может действовать как контролирующий элемент,
при этом скорость его секреции поддерживает определенные уровни показателей —
концентрацию глюкозы в крови, осмотическое давление крови или другой физиологический
параметр, который в норме сохраняется на постоянном уровне. Слежение за
системой осуществляется специфическими рецепторами, которые направляют
информацию об отклонениях регулируемого параметра от нужного значения в
центральный контроллер (от англ. controller — регулятор) системы. Контроллер
посылает сигнал в нервной или гормональной форме к эндокринной железе, в
результате чего ее секреторная активность снижается или возрастает. Латентный
период действия разных гормонов может составлять минуты и часы. В этих случаях
постоянный уровень концентрации гормона необходим для правильного осуществления
различных функций (пермиссивное действие).
2.2.4. Механизм действия гормонов
Действие гормонов основано на стимуляции или угнетении
каталитической функции некоторых ферментов в клетках органов—мишеней. Это
действие может достигаться путем активации или ингибирования уже имеющихся
ферментов. Причем важная роль принадлежит циклическому аденозинмонофосфату (цАМФ),
который является здесь вторичным посредником (роль первичного
|
|
|
Рис. 2.3 Механизм действия
гормонов — белковых, полипептидных и производных аминокислот |
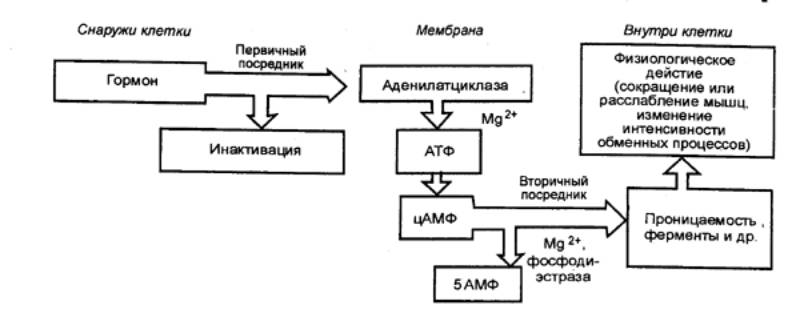
посредника выполняет сам гормон).
Возможно также увеличение концентрации ферментов за счет ускорения их биосинтеза
путем активации генов.
Механизм действия гормонов пептидной
и стероидной природы
различен. Амины и пептидные гормоны не проникают внутрь клетки, а
присоединяются на ее поверхности к специфическим рецепторам в клеточной
мембране. Рецептор связан с ферментом аденилатциклазой. Комплекс гормона
с рецептором активирует аденилатциклазу, которая расщепляет АТФ с образованием
цАМФ. Действие цАМФ реализуется через сложную цепь реакций, ведущую к активации
определенных ферментов путем их фосфорилирования, они и осуществляют конечный
эффект гормона (рис. 2.3).
|
|
|
Рис. 2.4 Механизм действия стероидных гормонов I — гормон проникает в клетку и связывается с рецептором в
цитоплазме; II — рецептор транспортирует гормон в ядро; III — гормон обратимо взаимодействует
с ДНК хромосом; IV — гормон активирует ген, на котором образуется матричная
(информационная) РНК (мРНК); V— мРНК выходит из ядра и инициирует синтез
белка (обычно фермента) на рибосомах; фермент реализует конечный гормональный
эффект; 1 — клеточная мембрана, 2 — гормон, 3 — рецептор, 4 — ядерная
мембрана, 5 — ДНК, 6 — мРНК, 7 — рибосома, 8 — синтез белка (фермента). |
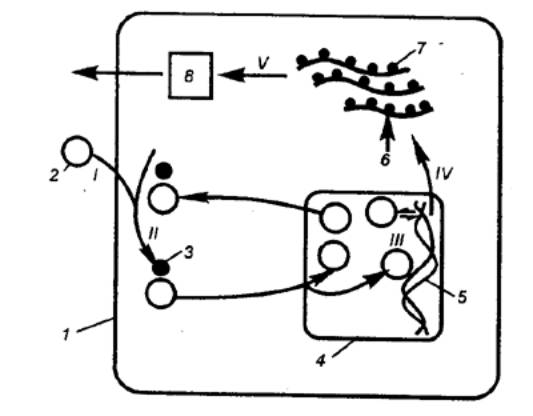
Стероидные гормоны, а также Тз и Т4 (тироксин и
трийодтиронин) растворимы в жирах, поэтому они проникают через клеточную
мембрану. Гормон связывается с рецептором в цитоплазме. Образовавшийся гормон—рецепторный
комплекс транспортируется в ядро клетки, где вступает в обратимое
взаимодействие с ДНК и индуцирует синтез белка (фермента) или нескольких
белков. Путем включения специфических генов на определенном участке ДНК одной
из хромосом синтезируется матричная (информационная) РНК (мРНК), которая
переходит из ядра в цитоплазму, присоединяется к рибосомам и индуцирует здесь
синтез белка (рис. 2.4).
В отличие от пептидов, активирующих
ферменты, стероидные гормоны вызывают синтез новых ферментных молекул. В связи
с этим эффекты стероидных гормонов проявляются намного медленнее, чем действие
пептидных гормонов, но длятся обычно дольше.
На основании функциональных критериев различают три
группы гормонов: 1) гормоны, которые оказывают влияние непосредственно на
орган—мишень; эти гормоны называются эффекторнымщ 2) гормоны, основной
функцией которых является регуляция синтеза и выделения эффекторных гормонов;
эти гормоны называют тропнымщ
3) гормоны, вырабатываемые нервными клетками и регулирующие синтез и
выделение гормонов аденогипофиза; эти гормоны называются рилизинг—гормонами,
или либеринами, если они стимулируют эти процессы, или ингибирующими гормонами,
статинами, если они обладают противоположным действием. Тесная связь между ЦНС
и эндокринной системой осуществляется в основном с помощью этих гормонов.
В сложной системе гормональной регуляции организма различают
более или менее длинные цепи регуляции. Основная линия взаимодействий: ЦНС →
гипоталамус → гипофиз → периферические эндокринные железы. Все
элементы этой системы объединены обратными связями. Функция части эндокринных
желез не находится под регулирующим влиянием гормонов аденогипофиза (например,
паращитовидные железы, поджелудочная железа и др.).
2.3. ЕДИНСТВО НЕРВНЫХ И ГУМОРАЛЬНЫХ МЕХАНИЗМОВ РЕГУЛЯЦИИ
Нейрогуморальная регуляция функций обеспечивает
адаптационные реакции от субклеточных до поведенческих. Можно выделить два вида
механизмов, лежащих в ее основе: жесткие (фиксированные) и гибкие
(нефиксированные). В основе работы жестких механизмов регуляции лежат
генетические программы, закрепленные в процессе эволюции, фенотипические
влияния сказываются лишь на конкретных формах реализации этих программ.
Гибкие механизмы обеспечивают исполнение организмом сиюминутных задач и тут
же перестают функционировать. Например, регуляция функционирования дыхательного
центра (чередования вдоха и выдоха) определяется генотипическими механизмами,
фенотипически же могут меняться длительность каждой фазы и амплитуда этих
процессов в соответствии с сиюминутными потребностями организма.
Нефиксированные механизмы нейрогуморальной регуляции функций осуществляются
срочно создающимися нейронными ансамблями. При этом главным принципом
объединения нейронов является доминанта, которая обеспечивает синхронизацию
работы входящих в ансамбли нервных образований. Количество, функциональная и
структурная принадлежность нейронов, входящих в это объединение, определяются
конкретными задачами регуляции, а также кинетикой формирования и осуществления
программы.
Вычленяют три вида регулирующих влияний: пусковые,
вызывающие деятельность регулируемой структуры или прекращающие ее; адаптационные,
влияющие на выраженность реакции и соотношение отдельных ее компонентов при
осуществлении функции; наконец, так называемые влияния готовности, формирующие
состояние готовности регулируемых образований.
Составная часть рассматриваемой проблемы — нейротрофические
влияния. В настоящее время общепризнано, что трофическая функция
свойственна всем нервным проводникам. Присуща она и симпатическим нервам в
отношении утомленной скелетной мышцы. Однако ее реализация происходит не
вследствие прямого действия этих нервов на мышцу, а опосредованно через систему
кровообращения. В исследованиях механизмов трофических влияний двигательных
нервов на поперечнополосатую мышцу удалось показать, что хотя импульсные
(медиаторные) и неимпульсные влияния тесно связаны, ведущее значение имеют
последние. Однако химическая природа нейрофических соединений (трофогенов),
приносимых к мышце аксоплазматическим транспортом из сомы, неясна.
В качестве нейросекреторной клетки можно рассматривать и
моторный нейрон, который передает мышечной клетке долгосрочную информацию,
необходимую для поддержания ее структуры и функции. Факт, что многие свойства
мышечных клеток регулируют посредством этого механизма, был подтвержден
перестройкой различных свойств быстрых и медленных мышц, происходящей после
перекрестной реиннервации. Это было показано прежде всего для сократительных
свойств мышц, которые изменились в соответствии с новой, «чужой» иннервацией.
Можно предполагать, что аналогичные общие закономерности существуют также и в
межнейронных отношениях. Эта точка зрения подкрепляется сведениями о
последующих событиях в нейронах после аксотомии (перерезки аксонов)
конвергирующих на них нервов.
Участие нейротрофических механизмов предполагают при нервных
травмах и восстановлении после них, в компенсаторных реакциях нервной системы,
в процессах памяти и пластичности, в период старения и при некоторых нервных и
мышечных заболеваниях. Ряд заключений о причинах нервной травмы был сделан в
опытах с временным прекращением проводимости, а также плазматического тока
(посредством действия цитостатика) при сохранении целостности аксонов. После
таких повреждений был сделан вывод, что анатомическая непрерывность нерва, а не
импульсная активность препятствует атрофии и фибрилляции мышцы.
2.3.1. Саморегуляция функций
организма
Саморегуляция физиологических функций — основной механизм поддержания
жизнедеятельности организма на относительно постоянном уровне. Саморегуляция,
возникнув в процессе эволюции как результат приспособления к воздействиям
окружающей среды, присуща всем формам жизнедеятельности. В ходе естественного
отбора в процессе приспособления к среде организмами были выработаны общие
регуляторные механизмы различной физиологической природы (нейрогуморальные,
эндокринные, иммунологические и др.), направленные на достижение и поддержание
относительного постоянства внутренней среды.
Способность поддерживать относительное постоянство
внутренней среды появляется на сравнительно высоких ступенях развития мира
животных. Так, уже у хрящевых рыб концентрация солей в крови и тканях
независима от ее изменения во внешней водной среде. У ганоидных и костистых рыб
также поддерживаются многие константы внутренней среды.
У человека и высокоорганизованных
животных гомеостатические
механизмы достигли высокой степени
развития. Относительное постоянство внутренней среды у них поддерживается
нервно—гуморальными физиологическими механизмами, регулирующими деятельность
сердечно—сосудистой и дыхательной систем, желудочно—кишечного тракта, почек и
потовых желез, которые обеспечивают удаление из организма продуктов обмена
веществ.
К наиболее совершенным гомеостатическим механизмам у высших
животных и человека относятся процессы терморегуляции. Определенные
признаки терморегуляции наблюдаются уже у животных с непостоянной температурой
тела, пойкилотермных (холоднокровных) животных, температура тела
которых в большинстве случаев следует за изменениями температуры внешней среды
не совсем пассивно. Однако о действительной терморегуляции можно говорить
только по отношению к гомойотермным (теплокровным) животным. Постоянство
температуры тела настолько велико, что в норме ее отклонение в «ядре тела» не
превышает нескольких десятых градуса при самых резких колебаниях температуры
внешней среды. В поддержании нормальной температуры тела участвует большое
число сложных процессов регуляции, часть которых в настоящее время изучена.
Особое значение для организма имеет постоянство состава
крови. Хорошо известна устойчивость ее активной реакции (рН), осмотического
давления, содержания глюкозы, соотношения электролитов (натрия, калия, кальция,
хлора, магния, фосфора) и т. д. Например, рН крови, как правило, не выходит за
пределы 7,35—7,47. Даже резкие нарушения кислотно—щелочного обмена с
патологическим накоплением кислот в тканевой жидкости, например, при
диабетическом ацидозе, очень мало сказываются на активной реакции крови.
Множество отдельных механизмов, регулирующих внутри— и
внесистемные взаимоотношения, оказывает в ряде случаев взаимопротивоположные
(антагонистические) воздействия, уравновешивающие друг друга. Это приводит к
установлению подвижного физиологического фона и позволяет живой системе
поддерживать относительное динамическое постоянство, несмотря на
изменения в окружающей среде и сдвиги, возникающие в процессе жизнедеятельности
организма.
Гомеостаз и гомеостатическая регуляция. Среди целостных реакций организма,
определяющих само его существование, поддержанию постоянства внутренней среды
принадлежит особая роль.
Понятие гомеостаз (от греч. homoios — подобный, схожий, и stasis — стояние, неподвижность)
(гомеостазис) впервые было введено в 1929 г. Уолтером Кенноном, творчески
развившим идеи К. Бернара о динамическом характере постоянства внутренней
среды. Этот термин У. Кеннон употреблял в двух смыслах: как постоянство
внутренней среды организма, обеспечиваемое деятельностью ряда физиологических
процессов, и как совокупность последних. Современные определения гомеостаза
включают оба аспекта.
Под внутренней средой У. Кеннон понимал кровь, лимфу и
тканевую жидкость. Основные параметры, характеризующие внутреннюю среду, были
названы гомеостатическими константами. В качестве примера назовем
концентрацию глюкозы, натрия и других ионов, величину мембранного потенциала,
значения артериального и осмотического давления, напряжения газов крови,
температуры. Они различаются по диапазону варьирования, т. е. норме реакции,
определяемой генотипом. Чем уже диапазон варьирования константы, тем более
значимы ее изменения для гомеостаза и тем большее число физиологических систем
участвует в ее регуляции. Примером могут послужить рН жидкостных сред и
температура тела у теплокровных. Гомеостатические константы с более широкой
нормой реакции расширяют адаптивные возможности организма. В каждый данный
момент гомеостатическая регуляция направлена преимущественно на достижение
оптимального уровня той константы, которая максимально отклонилась от своего
среднего значения.
Границы гомеостаза могут быть
жесткими и пластичными, меняться в зависимости от индивидуальных, возрастных,
половых, социальных и других условий. Жесткие константы, (например,
осмотическое давление крови) допускают лишь незначительные отклонения от своего
уровня, пластичные константы (например, уровень кровяного давления или
питательных веществ в крови) варьируют в довольно большом диапазоне и в течение
длительного времени. Значительная вариабельность уровня кровяного давления,
свойственная здоровому человеку в норме, имеет определенный физиологический
смысл. Так, повысившееся кровяное давление в результате физической нагрузки или
эмоционального сдвига улучшает кровоснабжение многих органов и тканей. Вместе с
тем длительное повышение кровяного давления приводит к нарушениям
кровоснабжения — кровоизлияниям, таким как инфаркты и инсульты.
Любые физиологические, физические,
химические или эмоциональные воздействия (будь то температура воздуха,
изменение атмосферного давления, обычная терапевтическая процедура или
волнение, радость, печаль, горе, прием лекарства и т. д.) могут явиться поводом
к выходу организма из состояния динамического равновесия, в котором он
пребывает. Таким образом, любое воздействие может оказаться «отклоняющим», или
«возмущающим». Рассмотрим достаточно простой пример. Углеводы служат важнейшим
источником энергии для организма. В результате распада и главным образом
«сгорания» в кислороде молекулы углеводов, богатые энергией, постепенно
превращаются в молекулы конечных продуктов — воды и двуокиси углерода,
обладающих малым запасом энергии. Энергия, высвобождающаяся при этом, идет на
покрытие энергетических потребностей клеток организма. Ни одна клетка, ни один
орган не могут существовать даже кратковременно без расходования энергии и
потребления «горючего» в виде углеводов.
Наиболее чувствительны к недостатку
снабжения «горючим» нервные и мышечные клетки. Особенно нервные, так как они
обладают незначительными запасами в виде гликогена и даже малое и
кратковременное снижение уровня сахара в крови (гипогликемия) приводит к
тяжелым функциональным расстройствам, вызывающим угрожающие явления в состоянии
всего организма. Функции нервных образований всецело зависят от содержания
сахара в крови. В крови здорового человека находится 44,4—66,6 мМ сахара в виде
глюкозы.
Обращает внимание строгое
постоянство уровня сахара в крови, что, по—видимому, наиболее благоприятно
для протекания процессов жизнедеятельности и обмена веществ. Оно обеспечивается
благодаря очень точно поддерживаемому балансу между потреблением сахара и его
поступлением в кровь. Существует не менее семи—восьми механизмов,
поддерживающих этот баланс. Центральную роль здесь играет печень.
Потребление сахара крови особенно
возрастает при повышенной мышечной деятельности. Можно было бы ожидать, что при
этом уровень сахара в крови резко понизится и наступит опасное состояние,
называемое гипогликемией. Однако этого не происходит: в печени как в
депо углеводов гликоген распадается до стадии глюкозы, которая и обеспечивает
замену сахара в крови. Можно было бы ожидать также и контрастного явления:
после приема пищи, богатой углеводами, последние, всасываясь в тонкой кишке, в
большом количестве поступают в кровь, что должно было бы привести к
значительному и стойкому повышению уровня сахара в крови. Но и этого не
наблюдается.
Это обусловлено тем фактом, что
оттекающая от кишки, обогащенная сахаром кровь поступает в общий кровоток не
сразу, а проходит сначала по воротной вене через печень. В клетках печени
глюкоза венозной крови поглощается, образуется гликоген, так что содержание
сахара в крови, поступающего из печени в общий кровоток, сохраняется
приблизительно на нормальном уровне. При употреблении очень большого количества
сахара наблюдается лишь небольшое и кратковременное увеличение содержания его в
крови, так называемая алиментарная гипергликемия. В этой ситуации
вследствие превышения «почечного порога» для глюкозы ее избыток удаляется с
мочой.
Какие же адекватные раздражители
возбуждают и приводят в действие механизмы регулирования уровня сахара в
крови? Последние могут, очевидно, вступать в действие только в ответ на
определенные возмущающие стимулы. Можно с уверенностью утверждать, что таким стимулом
является сам уровень сахара в крови, изменения которого обусловливают и
определяют регуляторную деятельность организма. Эта деятельность осуществляется
в тех случаях, когда уровень сахара в крови становится выше или ниже нормы.
Вступление в действие регуляторных механизмов вызывается изменением той
величины, постоянство и регулирование которой необходимы организму. Этот факт
свидетельствует о наличии здесь обратной связи, аналогичной техническому
замкнутому контуру регулирования. В самом деле, изменение уровня сахар в крови
вызывает противоположные возмущению регулирующие действия. Последние вызывают
новые изменения уровня сахара крови; их сдвиги приводят в свою очередь к новым
поправкам и т. д.
Применяя техническую терминологию, можно
подчеркнуть, что цепь регулирования уровня сахара в крови представляет собой
«систему с переходным и передаточным запаздыванием». Необходимым условием
всякого регулирования является непрерывное измерение подлежащей регулированию
величины с помощью рецепторного измерительного устройства. Полагают, что
рецепторы периферии (печени, поджелудочной железы), как и сахарочувствительные
клетки центров (гипоталамуса), формируют афферентный поток сигналов,
преобразуемый в промежуточном мозге и гипофизе в эфферентные посылы, исходящие
из центров и несущие приказы исполнительным механизмам. Исключительно важную
роль в регуляции уровня сахара в крови играют гормоны коры надпочечников (глюкокортикоиды),
их мозгового вещества (адреналин), а также поджелудочной железы (инсулин
и глюкагон) и щитовидной железы (тироксин) (см. гл. 6).
Закончив краткое рассмотрение
углеводной регуляции, вернемся снова к фундаментальному физиологическому закону
— гомеостазу. Любое раздражение, особенно стресс, ведет к
возникновению сложного комплекса реакций, основная цель которых приспособить
организм к изменившимся условиям, предотвратить или сгладить возможный сдвиг во
внутренней среде, в состоянии и деятельности органов, физиологических систем,
организма в целом.
Для развития организма постоянно
необходимо дополнительное количество энергии и пластических веществ, которое не
может приобрести полностью уравновешенная гомеостатическая система. Иначе для
реализации программы развития организма необходимо нарушение стабильности,
нарушение гомеостаза. Таким образом, наряду с законом сохранения гомеостаза в
развивающейся системе должен соблюдаться и другой закон — закон отклонения
гомеостаза.
Законы гомеостатической регуляции. Изменения гомеостатических констант
под влиянием воздействующих факторов должны устраняться системой
механизмов гомеостатического
регулирования. Закономерности самих изменений гомеостатических констант
описываются несколькими правилами.
Выведенное на основе экспериментальной и клинической
практики правило фона гласит: «Направленность и величина изменения
гомеостатической константы под влиянием воздействующего фактора зависят от ее
исходных (фоновых) значений». В качестве примера можно привести изменения
терморегуляции у человека, выходящего из холодного помещения (+12,6 °С), где он
озяб, на улицу (—15,6°С) и вновь возвращающегося в ту же комнату. При выходе
человека на улицу терморегуляция уменьшается теплоотдача за счет
вазоконстрикции кожных сосудов, усиливаются сократительный термогенез,
катаболические процессы в печени, пищеварительном тракте, увеличивается просвет
сосудов во внутренних органах возрастает тонус скелетных и гладких мышц. На
фоне увеличенного термогенеза и уменьшенной теплоотдачи, адаптированных к —15,6°С,
возврат в помещение (+12,6 °С) вызывает первоначальное ощущение перегрева и
способствует запуску реакций противоположной направленности с последующей
адаптацией терморегуляции к новому температурному режиму окружающей среды.
Следовательно, гомеостатическая регуляция, направленная на стабилизацию
температуры тела человека, меняется и в зависимости от фона может
осуществляться разными механизмами.
Другая особенность гомеостатической регуляции сформулирована
в виде правила гиперкомпенсации: «Регуляция сдвигов гомеостатических
констант носит гиперкомпенсаторный характер». Говоря иными словами, новое
значение гомеостатической константы, достигнутое в результате
гомеостатирования, не идентично фоновому, а превышает его. Это соответствует
определению гомеостаза как динамического относительного постоянства внутренней
среды.
На клеточном уровне закон гиперкомпенсации можно
иллюстрировать изменениями мембранного потенциала возбудимой клетки: на смену
спайку с характерными для него деполяризационным сдвигом мембранного потенциала
и защелачиванием цитоплазмы приходит следовая гиперполяризация. Она
представляет собой гиперкомпенсаторные сдвиги мембранного потенциала (более
отрицательные значения, чем у потенциала покоя) и рН примембранной цитоплазмы
(ацидоз превышает фоновый).
Типы гомеостатической регуляции. Различаются по условиям запуска
соответствующих механизмов. Так, в приведенных выше примерах имеет место
гомеостатическая регуляция по отклонению, когда само изменение величины
константы вызывает запуск гомеостатических механизмов регулирования. Этот тип
регуляции характерен для тех случаев, когда воздействующий фактор является
новым для организма. По мере повторения воздействия и запоминания его
параметров наблюдается появление гиперкомпенсаторых изменений гомеостатических
констант, опережающих их первичные сдвиги. Подобная опережающая
гомеостатическая регуляция имеет энергосберегающее значение.
Опережающая гомеостатическая регуляция может стать причиной
извращения знака первичной реакции. Например, было показано, что употребление
алкоголя впервые вызывает у человека первичную реакцию снижения температуры
тела (регуляция по отклонению), тогда как при повторных применениях — ее
повышение (опережающая регуляция). Аналогично первичный электрошок
вызывает .тахикардию, а при повторных применениях — брадикардию. Этот же тип
опережающей гомеостатической регуляции лежит в основе повышения толерантности к
фармакологическим агентам при их
многократных введениях. Он используется при обучении пациентов и
экспериментальных животных регулированию собственного системного артериального
давления.
Не всегда опережающая
гомеостатическая регуляция связана с обучением в онтогенезе. Генетически
закрепленная память способствует включению опережающего гомеостатирования до
наступления запредельных или близких к ним сдвигов констант. Это видно на примере
поискового пищевого поведения (гомеостатирующее поведение по А. Д.
Слониму), которое запускается прежде, чем уровень глюкозы в крови уменьшится до
критических значений. В контроле опережающего гомеостатирования, связанного с
врожденной и приобретенной памятью, значительная роль принадлежит циркадным
ритмам активности нейроэндокринной системы организма, определяемым
соотношением периодов света и темноты, магнитным полем Земли, периодами
солнечной активности, лунными циклами и др.
На разных уровнях реализации
механизмов гомеостатирования соотношение между гомеостатической регуляцией двух
типов может различаться. Так, на субклеточном и клеточном уровнях преобладает
регуляция по отклонению, обусловливая последовательную смену реакций
фосфорилирования — дефосфорилированием, катаболизма — анаболизмом, эндоцитоза —
экзоцитозом, открытого состояния ионного канала закрытым и т. п. В целом
обеспечивается гомеостатирование внутриклеточного рН, осмотического давления и
объема клетки. На уровнях системной регуляции значений основных
гомеостатических констант оба типа регуляции равноправны, тогда как на уровне
организма преобладает опережающая. Все уровни регуляции взаимосвязаны, причем
их иерархия определяется порогом чувствительности к изменениям регулируемой
гомеостатической константы и возможностями ее регуляции на данном уровне.
Для большинства уровней регуляции
значений каждой константы характерна взаимозамещаемость гомеостатических
механизмов и многоконтурность регуляторных воздействий. Так, умение
регулировать уровень собственного артериального давления при гипотонии можно
выработать несколькими способами: внушением, воспоминанием об эмоционально
напряженной ситуации, повышением тонуса определенных скелетных мышц или
специальными упражнениями, а также задержкой дыхания. С каждым из них связаны
различные нейрогормональные механизмы регуляции артериального давления,
действующие на разные компоненты сердечно—сосудистой системы и контролирующие
ее центры через запуск соответствующих реакций. Этим достигается надежность и
гибкость гомеостатической регуляции значений данной константы. Следует
подчеркнуть, что в живом организме никогда не происходит селективной регуляции
какой—либо одной константы, поскольку константы взаимосвязаны. Она приводит к
минимизации энергетических затрат на решение таких комплексных задач, как
выработка и сохранение оптимального режима взаимодействия физиологических
систем или организма со средой в изменившихся условиях, поддержание
определенных уровней стационарного состояния и сохранение целостности
организма.
Закрепившееся в процессе
эволюционного развития состояние гомеостаза позволяет организму
приспосабливаться к условиям окружающего мира. Адаптация при этом может быть
оптимальной, неоптимальной и даже вредной, связанной с нарушением жизнедеятельности.
Живая система способна перестраиваться, переходить на новый гомеостатический
уровень, активизируя при этом одни регулирующие системы и тормозя другие.
Адаптация к стрессорным факторам осуществляется на всех
уровнях организации начиная с клеточного, однако для реализации
гомеостатической защитной реакции у высших животных имеется специализированная адаптационная
система. Основными компонентами этой системы являются:
кора надпочечников, вырабатывающая гормон защиты — кортизол;
гипофиз, который высвобождает кортикотропин, регулирующий продукцию
кортизола;
и, наконец, гипоталамус,
контролирующий секрецию кортикотропина, а также другие отделы ЦНС (см. разд.
6.2).
2.3.2. Обратная связь как один из
ведущих механизмов в регуляции функций организма
Процесс саморегуляции постоянно сохраняет циклический
характер и осуществляется на основе правила: какое—либо отклонение от
константного уровня любого жизненного фактора служит толчком к немедленной
мобилизации аппаратов, вновь восстанавливающих этот постоянный уровень.
По своей природе физиологическая саморегуляция является
автоматическим процессом. Факторы, отклоняющие константу, и силы, ее
восстанавливающие, всегда находятся в определенных количественных соотношениях.
В этом физиологическая саморегуляция тесно соприкасается с закономерностями,
сформулированными кибернетикой, теоретическим стержнем которой является
автоматическая регуляция заданного фактора при помощи замкнутого контура с
обратной связью. Наличие обратной связи уменьшает влияние изменений параметров
системы на работу ее в целом, обеспечивает также ее стабилизацию и
устойчивость, улучшает переходные процессы, обеспечивает повышение ее
помехоустойчивости.
|
|
|
Рис. 2.5 Гомеостатическая система управления. Стрелками показаны направления
воздействий. |
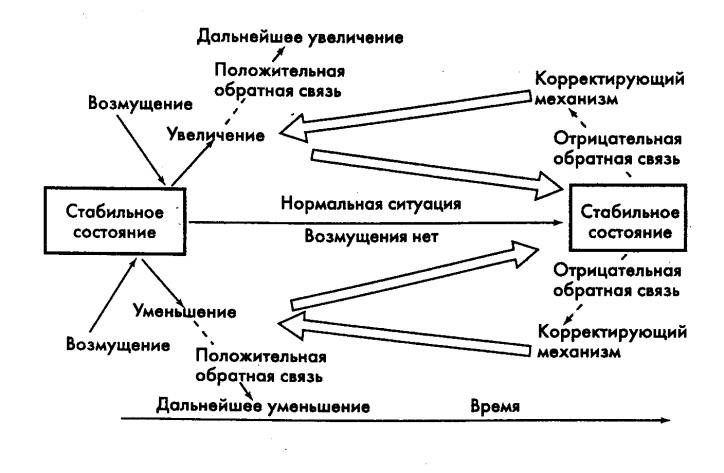
Как уже указывалось, связь выхода системы с ее входом через
усилительное звено с положительным коэффициентом усиления — положительная
обратная связь, с отрицательным усилением — отрицательная обратная связь
(рис. 2.5). Положительная обратная связь повышает коэффициент усиления и
обеспечивает возможность управления значительными потоками энергии, затрачивая
при этом малые энергетические ресурсы. Заметим, однако, что в биологических
системах положительная обратная связь реализуется в основном в патологических
ситуациях. Отрицательная обратная связь обычно улучшает устойчивость
системы, т. е. ее свойство возвращаться к первоначальному состоянию после
прекращения влияния внешнего возмущения. Требование устойчивости — одно из
основных требований для системы управления, так как устойчивость определяет,
как правило, работоспособность всей системы.
Обратные связи в организме обычно иерархичны,
накладываются одна на другую и дублируют друг друга. Их можно делить по разным
категориям, например, по постоянной времени — на быстродействующую нервную и
более медленную гуморальную и т. д. Например, ту же систему регулирования
сахара в крови следует рассматривать как многоконтурную.
В 30—е гг. советский биолог М. М.
Завадовский на основании изучения гуморальных механизмов регуляции в растущем
организме выдвинул общебиологический принцип регуляции процессов развития и
гомеостаза «плюс — минус взаимодействие». Сущность этой концепции сводится к
следующему. Если между двумя органами (процессами) имеется непосредственная
связь, причем первый орган (процесс) стимулирует второй, то второй тормозит
первый, и наоборот. В сущности, речь идет о механизме обратной связи. При этом
прямая и обратная связи между органами и процессами имеют противоположные
знаки: плюс—минус, минус—плюс. Такого типа связи обеспечивают животному и
человеку свойства саморегулирующейся системы с высокой степенью устойчивости.
В ходе исследования роли афферентной
информации в осуществлении локомоторных актов Н. А. Бернштейн выдвинул идею о сенсорных
коррекциях, в соответствии с которой непрерывное участие потока афферентной
сигнализации контрольного или коррекционного значения является необходимым
компонентом двигательных реакций. Каждый случай упорядоченного реагирования
представляет собой непрерывный циклический процесс взаимодействия организма с
меняющимися условиями окружающей или внутренней среды. При этом огромную роль
играет контролирующая коррекционная афферентация.
Другой советский физиолог, П. К.
Анохин, еще в 30—е гг. и, пожалуй, впервые четко обосновал понятие обратной,
или санкционирующей, афферентации, т. е. обязательной при любом действии
импульсации, которая идет от рецепторов в ЦНС и информирует о результатах
произведенного действия, соответствующего или не соответствующего намеченной
цели. При дальнейшей разработке механизма последний получил название акцептора
результата действия.
Примеров осуществления обратных
связей в организме множество. Рассмотрим лишь некоторые процессы регулирования
в нервной системе. Распространение нервных влияний отдаленно напоминает
железнодорожное движение от одной станции до другой. Грузооборот станции
определяется в основном не ее величиной, количеством помещений и т. д., а
густотой и пропускной способностью ее линий связи с другими станциями.
Аналогичным образом и в нервной системе упор в регуляции зачастую делается на
доклеточное звено — синаптический аппарат. Подобно семафорам и стрелкам, перед
которыми часто останавливается движение, в нервной системе осуществляется пресинаптическое
регулирование. Суть его заключается в том, что импульсы возбуждения,
бегущие по одним волокнам, благодаря специализированному промежуточному нейрону
затрудняют распространение таких же импульсов по другим нервным волокнам и
«поезд останавливается перед семафором».
В центральной нервной системе
существует еще один вид регуляции, пожалуй, наиболее изученный, осуществляемый
на выходе рефлекторной дуги, —
возвратное торможение. В этом случае импульсы,
распространяющиеся от моторной клетки к мышцам, частично возвращаются в спинной
мозг и через специализированный промежуточный нейрон — клетку Реншоу —
уменьшают активность тех же или других моторных нейронов, десинхронизируя их
деятельность. В результате этого мышечные волокна сокращаются не одновременно,
что обеспечивает плавность мышечных движений. Пример с мотонейронами спинного
мозга, возможно, самый яркий, но вообще подобные способы саморегуляции
рефлекторной деятельности по типу отрицательной обратной связи широко
распространены в ЦНС.
Значение механизмов обратной связи в поддержании гомеостаза
чрезвычайно велико. Так, поддержание константного уровня кровяного давления
всегда является следствием взаимодействия двух сил: нарушающей этот уровень и
восстанавливающей его. В результате увеличенной импульсации из барорецептивных
областей (главным образом синокаротидной зоны) снижается тонус вазомоторных
симпатических нервов и повышенное кровяное давление нормализуется. Депрессорные
реакции в норме оказываются сильнее прессорных реакций.
Увеличение содержания в крови катехоламинов — адреналина и
норадреналина — при их инъекции или при естественной реакции организма на
внешнее воздействие приводит к активации периферических эффекторных
образований, тем самым имитируя возбуждение симпатического отдела нервной системы,
но при этом препятствует дальнейшему высвобождению и синтезу таких соединений.
2.3.3. Рефлекторный принцип регуляции функций
В основе представлений о
нейрогуморальной регуляции функций лежит учение о рефлексе. Рефлексом
называют любую ответную реакцию организма, осуществляющуюся с участием нервной
системы.
Структурной основой осуществления
таких реакций является рефлекторная дуга, включающая пять звеньев: рецепторы,
воспринимающие воздействие раздражителей; афферентные нейроны,
связывающие своими отростками рецепторы с центрами нервной системы и
обеспечивающие центростремительное проведение сигнала; промежуточные,
или вставочные, нейроны, представляющие собой центральную часть рефлекторной
дуги, или нервный центр; эфферентные нейроны, по аксонам которых
происходит центробежное проведение сигналов к периферическим органам и тканям: эффекторы,
или исполнительные органы (поперечнополосатые скелетные или гладкие
мышцы внутренних органов, железы), осуществляющие соответствующую рефлекторную
деятельность.
Во всех учебниках утверждается, что
замыкательное звено рефлекторной цепи — центральная нервная система. Это в
подавляющем числе случаев так, однако в недавнем прошлом в ряде отечественных и
зарубежных лабораторий описаны и местные рефлексы, замыкающиеся в
периферических автономных ганглиях. Именно они во многом восстанавливают и
обеспечивают функции децентрализованных или пересаженных внутренних органов.
Разбирая структуру рефлекторной
дуги, надо иметь в виду, что она является замкнутой. Поступая в кровь,
физиологически активные вещества (прежде всего медиаторы и гормоны) составляют
в определенных условиях гуморальное звено рефлекторной дуги. Они передают в
спинной и головной мозг соответствующую
информацию, под влиянием которой возникает поток нервных импульсов из ЦНС в
эффекторные органы.
Классическая рефлекторная дуга, таким образом, усложняется,
превращаясь в многозвенное кольцо, в котором нервное звено сменяется
гуморальным, а гуморальное — .нервным.
Определенные рефлекторные реакции могут включать различное
количество рецепторов, афферентных и эфферентных нейронов и сложные процессы
взаимодействия возбудительных и тормозных процессов в центрах нервной системы.
Области тела, в пределах которых расположены специализированные рецепторы,
раздражение которых обусловливает строго определенные рефлексы, получили
название рефлексогенных зон, или рецептивных полей.
Афферентный путь проведения информации от рецепторов в ЦНС обычно включает в
себя биполярный нейрон спинальных ганглиев. Помимо этого пути чувствительными
проводниками могут служить вегетативные нейроны — клетки Догеля II типа,
способные образовывать релейные соединения на различных уровнях нервной
системы.
В отличие от редких случаев, отмеченных для соматических
нервных образований (регуляция деятельности скелетных мышц), для всех
вегетативных рефлексов характерна полисинаптичность рефлекторной дуги.
Структурная и функциональная организация нервных центров наиболее изучена на
уровне спинного и продолговатого мозга. Надсегментарные рефлекторные процессы,
довлеющие над ними и вовлекающие миллионы и миллиарды нейронов разной
морфологической, физиологической и химической природы, изучены гораздо хуже.
Необходимо учитывать, что нервные импульсы при всяком рефлекторном акте
способны широко распространяться в центральной нервной системе по
многочисленным проводящим путям, включая стохастически функционирующие
интернейроны.
Благодаря иррадиации возбуждения в ЦНС через один
афферентный вход воздействие может распространиться на значительное число
эфферентных нейронов. Взаимоотношения между рефлексами — содружественные и
антагонистические, их цепной характер — предмет особого подробного
рассмотрения. Не менее подробного освещения заслуживают и процессы интеграции,
синтеза разнородных по своей природе возбуждений в ЦНС.
Особое внимание должен привлечь феномен доминанты,
сформулированный академиком А. А. Ухтомским как временно господствующая
рефлекторная система, направляющая работу нервных центров в данный момент.
Интимные механизмы доминанты не определены, феноменология же ее вполне
конкретна. Предварительно подготовленное импульсами своей рефлекторной дуги,
подкрепленное в результате суммации влияниями из постороннего источника,
возникающее в доминантном очаге возбуждение тормозит прочие рефлексы на
очередные раздражители.
В развитии рефлекторной теории чрезвычайно важный вклад был
сделан И. М. Сеченовым, распространившим понятие рефлекторных актов на все
характеристики поведения человека, в том числе и на его психические
проявления. Однако упрощенное понимание рефлекса как механизма,
заканчивающегося унитарным рефлекторным действием, по существу «останавливало»
понимание динамики развития поведения организма и не полностью раскрывало
приспособительное значение рефлексов. Учение об условном рефлексе И. П.
Павлова позволило сделать существенный шаг вперед в развитии рефлекторной
теории и познании механизма рефлекторного действия. Ниже рассмотрены общие
понятия о системах.
Конкретным аппаратом саморегуляции функций организма
является функциональная система, представляющая обширное взаимодействие
центральных и периферических образований, составляющих совокупно действующий
комплекс с определенными физиологическими свойствами. Такой аппарат может
включать в себя различные анатомические образования, комбинации гуморальных
веществ, объединенных взаимозависимостью и соучастием в каких—либо
приспособительных реакциях организма.
Каждая функциональная система, по определению П. К. Анохина,
представляет собой до некоторой степени замкнутую систему благодаря тому, что
позднее назвали «обратной связью», осуществляемой с периферических
исполнительных органов. Таким образом, каждая функциональная система имеет
определенный комплекс афферентных посылов, который через акцептор действия
определяет выполнение функции. Так, при дыхательном акте афферентные импульсы
следуют от диафрагмы, трахеи, легких, межреберных мышц. Несмотря на их
различное происхождение, эти влияния интегрируются в центральной нервной
системе путем временных и тонких соотношений между ними.
В организме существуют и регуляторные системы,
работающие не по принципу рассогласования, а с учетом оценки величины
поступающего сигнала, который нарушает состояние системы. Толчком к их
деятельности служит отклонение от заданной величины не на выходе, а на входе
системы, т. е. действие на систему раздражителей, отличающихся от заданных
параметров. На входе этого регулятора «по возмущению» имеются устройства,
которые улавливают и оценивают величину поступающего сигнала, нарушающего
состояние системы. Если эта величина превышает допустимую и способна вызвать
нежелательные отклонения в состоянии системы, то возникают команды,
обеспечивающие нейтрализацию влияния этих сигналов и сохранение стабильного
состояния системы. В этой ситуации происходит не восстановление уже нарушенного
состояния системы, а предупреждение возможности таких нарушений.
Пример пресинаптического торможения в нервной системе
приводился, но надо иметь в виду, что оно является только частным проявлением
пресинаптического регулирования. Пресинаптические терминали с их богатым
рецепторным аппаратом во многих случаях являются ареной действия гуморальных
веществ, в результате чего и осуществляется коррекция синаптического процесса и
рефлекторной дуги в целом.
В регуляторных, защитных, компенсаторных — любых
физиологических реакциях имеет место взаимодействие обоих принципов и обоих
механизмов регуляции, функционирующих на входе и выходе системы.
Для любой функциональной системы необходимо получение информации
о ее состоянии, о величине поступающих сигналов и возникающих при этом сдвигах
в ее деятельности. Необходим аппарат сличения параметров этих сдвигов или
параметров поступающих сигналов с величиной наличных и нормальных для данной
системы параметров. 'Конечно, необходим аппарат, формирующий команды,
предотвращающие эти сдвиги. Действие этих команд осуществляется путем
нормализации уже возникших отклонений и предупреждением нежелательных эффектов
возмущающего входного сигнала посредством уменьшения силы сигнала, снижения
чувствительности системы к данному возмущающему действию или предотвращения
его.
Абсолютным условием саморегуляторных приспособлений
организма является
наличие следующих факторов, придающих функциональной системе определенную
направленность действия.
1. Достигаемый в результате
регуляции приспособительный эффект или константа жизненно необходимы для
организма. Такие жизненно важные константы определяются генотипически.
Функциональная особенность каждой саморегуляции зависит от того, насколько она
податлива действию внешних и внутренних факторов, ее отклоняющих. Пример
«жесткой» функциональной, системы (осмотическое давление крови) и «пластичной»
(уровень кровяного давления) уже приводился.
2. Саморегуляция — циклический, фазовый процесс, разыгрывающийся
на вполне конкретном аппарате структур и механизмов, составляющих в
совокупности функциональную систему. Все саморегуляторные приспособления
диктуются фактом отклонения конечного приспособительного эффекта или
несоответствия силы входного возмущающего сигнала потребностям системы.
3. Одним из обязательных условий саморегуляции является
информация о конечном приспособительном эффекте в центральные регулирующие
аппараты организма, так же как и нивелирование нежелательных или чрезмерных влияний
«на входе» системы.
4. В зависимости от широты охвата органов и систем типы
саморегуляции могут быть различными по характеру. Одни из них осуществляются с
помощью аппаратов функциональных систем, компоненты которых не выходят за
пределы организма и потому конечный приспособительный эффект достигается на
основе внутренних циклов организма. К этой категории регуляций относится уже
упоминавшийся контроль уровня сахара в крови.
Примером работы функциональных систем с обширным комплексом
внешних факторов является саморегуляция количества питательных веществ,
находящихся в кровяном русле. Они служат источником обмена веществ в тканях, и
потому их истощение является непрерывным процессом, отражающим трату энергии
организмом. Специфический рецепторный аппарат этой достаточно пластичной
константы, по—видимому, расположен в латеральной области гипоталамуса.
Уменьшение уровня питательных веществ ведет к активации гипоталамических
образований, вовлечению в реакцию подкорковых, и корковых центров, что
проявляется в возникновении объективного ощущения голода. Параллельно
организуются специфические поведенческие акты, которые в целом получили
название пищедобывательного поведения. После добычи пищи, ее переработки
в желудочно—кишечном тракте, всасывания в кровь, концентрация питательных
веществ в ней повышается и этим самым снижается возбужденное состояние пищевых
центров гипоталамуса.
5. В случае экстремального воздействия на организм
саморегулирующиеся системы формируют защитно—приспособительные акции, сохраняют
постоянство конечного приспособительного эффекта и в сумме подчиняются ряду
общих законов развития. Среди этих законов важнейшим является тот, который
определяет соотношение воздействующего и защитного эффектов. В условиях
нормального протекания саморегулирующихся процессов сила максимально возможного
защитного приспособления организма всегда должна быть большей, чем выраженность
максимально возможного отклонения данного приспособительного конечного эффекта
от константного уровня.
Например, как бы ни было высоко кровяное давление,
количество возникающих на периферии депрессорных влияний в сумме должно быть
более сильным, чем те факторы, которые отклоняют уровень кровяного давления. В
этом и состоит причина того, что в норме, несмотря на эпизодические подъемы кровяного
давления, оно непременно возвращается к исходному уровню, обеспечивая
оптимальное снабжение органов кровью.
Не менее важным свойством саморегулирующихся систем является
опережающая мобилизация тех нервных импульсаций, которые возникают в рецепторах
приспособительного эффекта. Мобилизация защитных приспособлений организма
происходит в значительно большей степени, чем нарастание отклоняющих факторов.
Отмеченная закономерность приводит к тому, что с каждым новым этапом нарастания
отклонения происходит несоответственно высокая мобилизация обратной информации
в центральную нервную систему. Способность саморегулирующихся функциональных
систем сопротивляться отклонениям с помощью прогрессирующей мобилизации
защитных приспособлений с повышающейся силой охраняет жизненно важные
константы, делает маловероятными отклонения от нормы.
2.4. ОБЩИЕ ЧЕРТЫ КОМПЕНСАТОРНО—ПРИСПОСОБИТЕЛЬНЫХ
РЕАКЦИЙ ОРГАНИЗМА
Компенсаторные реакции — первичные адаптивные сдвиги,
направленные на устранение или ослабление нарушенной структуры или функций
живых систем, вызванных повреждающими или экстремальными воздействиями среды.
Компенсаторные механизмы — динамичные, быстро возникающие физиологические
средства аварийного обеспечения, развивающиеся на различных уровнях — от
молекулярного до организма в целом. Эти механизмы мобилизуются при попадании
организма в неадекватные условия и постепенно затухают в ходе развития
адаптационного процесса.
В широком смысле компенсация нарушенных функций
(полная или частичная, завершенная или временная) всегда является
«физиологической мерой организма», направленной на восстановление его рабочих
констант. Оценивая физиологическое содержание всех видов нарушенных функций
организма (перенапряжение, болезнь, травма и т. д.), можно сделать заключение,
что компенсация во многом является универсальным процессом и отличается лишь в
отдельных случаях нарушения физиологических систем специфическими и
количественными признаками (быстротой включения компенсаторных механизмов,
широтой вовлекаемых резервных механизмов, степенью участия коры головного мозга
и пр.).
В дидактических целях иногда выделяют внутриклеточные,
тканевые, органные, системные и межсистемные Компенсаторные реакции. Однако в
целом организме эти процессы на более низком уровне организации зачастую развертываются
на основе ограниченного числа общих стандартных механизмов и интегрированы
межсистемными компенсаторными реакциями.
В организме, представляющем собой единую гомеостатическую
систему, в результате непрерывного нейрогуморального взаимодействия компонентов
этой системы формируется их функциональная сопряженность. В том случае, если
функциональный дефект выражен сильно и не может быть компенсирован на местном
уровне перестройкой активности поврежденной структурой, тогда компенсаторная
реакция приобретает генерализованный характер, охватывает многие системы
организма и на всех уровнях их организации. Она начинается с осуществления
срочных, а затем более долговременных компенсаторных реакций.
Механизмы компенсаций являются составной частью резервных и
защитных сил организма, формируют надежность живых систем, т. е. их способность
выполнять определенные функции (безотказную работу) в течение некоторого
времени. Надежность, как и определяющие ее факторы, совершенствуется в процессе
онтогенеза. На основе компенсаторных механизмов формируются условнорефлекторные
реакции на обстановку, а также функциональные системы, обеспечивающие организму
необходимую резистентность и целенаправленное поведение в неадекватных
условиях. Вместе со специфическими компенсаторно—приспособительными реакциями в
организме складываются и неспецифические. Осуществление компенсации нарушенных
функций, свойство надежности физиологических систем обеспечивается рядом
правил:
1. Правило исходного состояния
(фона) постулирует зависимость направленности и величины эффекта
регулирующего воздействия от исходного значения метаболизма и функции
регулирующей системы. Если перед действием регулирующего сигнала обмен веществ
и функция системы снижены, то регулятор вызывает максимальный эффект. И наоборот,
при активированном состоянии объекта в ответ на стимулирующий регуляторный
сигнал отмечается слабый эффект или его отсутствие.
2. Правило компенсаторной
клеточной регенерации и гиперплазии ткани представляет собой реализацию
общебиологической способности тканей к росту и диффёренцировке. Эта способность
зависит от соотношения ряда факторов, в частности, гормонов—регуляторов и
ингибиторов клеточного роста в микросреде клеток. К числу регуляторов относятся
инсулин, соматотропин, тиреоидные и, возможно, другие гормоны, а также
полипептидные ростовые факторы клеточного происхождения: фактор роста нервов,
эпидермиса, фибробластов, тромбоцитарный и трансформирующий факторы роста. В
реализации процессов регенерации и гиперплазии играют роль также и ростовые ингибиторы,
к числу которых относят простагландины, циклические нуклеотиды и кейлоны.
3. Правило избыточности. Он
связан с существованием гораздо большего, чем необходимо для осуществления
функции, числа элементов. Притом выявляется не только структурная, но и
функциональная информационная избыточность. В технических системах с
возрастанием количества элементов снижается надежность их функционирования. С
усложнением живых систем, напротив, надежность работы возрастает в связи с
улучшением механизмов их регулирования.
4. Правило дублирования в системах регулирования
проявляется не только присутствием в организме одинаковых структур, например,
парных органов (почки, легкие, надпочечники), но и существованием
многочисленных структур с одинаковой функцией (клетки разных тканей, нейроны
мозга, нефроны почек и др.). Особенно демонстративны примеры дублирования в
работе ЦНС. Экспериментальные данные свидетельствуют, что имеется выраженный
динамизм мобилизации определенных структур мозга при получении одного и того же
эффекта. Различные нейронные ансамбли ЦНС могут участвовать в реализации не
одного, а нескольких механизмов системной деятельности. Степень участия той или
иной ансамблевой структуры на каждом из этапов осуществления системных
процессов является различной, изменчивой по мере образования и закрепления
адаптивных навыков.
Способность мозга к компенсации нарушенных функций не
является жестко регламентированной, неизменной. Она предполагает достаточно
длительную перестройку предшествующих отношений между центром и периферией,
замену их новыми функциональными отношениями на основе длительной тренировки,
упражнения.
Вклад различных отделов мозга в компенсацию функций
организма не одинаков. Решающее значение в осуществлении приспособительных
перестроек имеет кора больших полушарий мозга. Если у животных, достигших
предела компенсации после предшествующих различных нарушений деятельности
внутренних органов, удалить кору больших полушарий, то у них наблюдается
вторичная длительная декомпенсация функций, приобретающая в ряде случаев
характер необратимых нарушений. Удаление высшего отдела мозга у животных с
последующим повреждением других отделов ЦНС превращало их в глубоких инвалидов,
и компенсации функций у них не наступало. Компенсаторные возможности изменяются
в ходе постнатального онтогенеза: у молодых животных при тех же видах
оперативного вмешательства, что и у взрослых, они намного выше.
Еще один пример: роль множества параллельно действующих
контуров регуляции висцеральных органов. Надежности работы, резистентности
внутренних органов при патологии способствуют особенности их сложной автономной
(вегетативной) иннервации, и прежде всего, наличие внутриорганных
метасимпатических рефлекторных дуг (см. главу 5). Установлены многозвенные и
многоэтажные цепные реакции, морфологическим субстратом осуществления которых
являются кольцевые взаимосвязи внутренних органов, сосудов, желез внутренней
секреции. Так, целая гамма периферических рефлексов (пусковых, стимулирующих и
тормозящих корригирующих) отмечена во взаимоотношениях кишки, желудка, жёлчного
пузыря, мочевыводящих путей.
5. Правило резервирования функций
связан с существованием в системе «дежурных» и «резервных» элементов, способных
переходить от покоя к деятельности. Чаще всего это происходит при
интенсификации функционирования. Например, для увеличения силы мышечного
сокращения мобилизуются дополнительные двигательные единицы из числа молчавших
α—мотонейронов спинного мозга. Вовлечение резервных элементов особенно
важно при повреждениях функциональной системы. Зачастую это обеспечивает
сохранение функции.
6. Правило периодичности
функционирования составляющих элементов определяет особенности поведения
системы в состоянии покоя. В тканях происходит смена открывающихся и закрытых капилляров,
в легких — вентилируемых альвеол, в мозговых центрах — активных нейронов и т.
д. Выявлена вероятность (стохастичность) нейрональной активности, особенно в
высших отделах мозга. Вариабельность реакций нейронов не является помехой в
деятельности ЦНС, она обеспечивает высокую функциональную надежность,
детерминизм работы центров, обусловленный вероятным способом участия нейронов в
ансамблях мозга. Таким образом, функциональными элементами центров ЦНС служат
не отдельные клетки, а нейронные ансамбли, реализующие свои функции
статистически, и участие отдельных нейронов в каждом таком ансамбле не
фиксированное, а в той или иной степени вероятностное. Именно это
обстоятельство обеспечивает большие регуляторные возможности ЦНС, ее
потенциальные способности к компенсации нарушенных функций.
7. Правило взаимозаменяемости и
смены функций обеспечивает возможность достижения компенсации при нарушении
одной из функций за счет активности другой. Это можно проиллюстрировать хорошо
изученной системой кардио—респираторного сопряжения. При затруднениях внешнего
дыхания усиливается работа сердечно—сосудистой системы, активируется
эритропоэз, и все это обеспечивает достаточную доставку кислорода тканям.
8. Правило усиления в системах регуляции и
прогрессивной мобилизации запасных приспособительных механизмов с
использованием многоконтурного воздействия на измененный параметр, что приводит
к минимизации энергетических затрат на приспособительные процессы.
Использование в живых системах незначительных сигналов для получения мощного
эффекта хорошо известно. Так, в системе гормональной регуляции используется
каскадный процесс:
либерины и статины гипоталамуса —
тропные гормоны гипофиза — гормоны рабочих желез.
9. Правило повышения чувствительности денервированных
(децентрализованных) структур проявляется в увеличении реактивности
образований, лишенных нервного влияния. По—видимому, это реализуется за счет
расселения субсинаптических рецепторов в более обширные области
постсинаптической мембраны, увеличения их числа и сродства к гуморальным
влияниям. Этот принцип проявляется в частичном восстановлении функций
децентрализованных внутренних органов, в частности, за счет усиления
замыкательных свойств внутри— и внеорганных автономных (вегетативных)
рефлекторных дуг. Повышенная чувствительность после децентрализации и
денервации, характерная для всех возбудимых тканей, представляет собой одно из
проявлений выпадения трофической функции нервной системы. Вероятно, то, что
называется сенситизацией, стабилизацией, или повышенной чувствительностью,
представляет собой истинную чувствительность клеток, обычно маскируемую
нервными влияниями.
2.4.1. Общий адаптационный синдром
Адаптация является совокупностью физиологических реакций,
обеспечивающих приспособление строения и функций организма или его органа к
изменению окружающей среды с включением стресс—реализующей системы (прежде
всего гипоталамо—гипофизарно—надпочечниковой и адреналовой). В
концентрированном виде концепция неспецифических реакций организма на сильное
воздействие нашла воплощение в понятии «общий адаптационный синдром» (Г.
Селье).
Прослеженные закономерности появления защитных реакций после
воздействия стрессорного фактора позволили выделить три фазы общего
адаптационного синдрома: реакцию тревоги — «аварийная фаза», переход к
устойчивой адаптации и повышенной резистентности (сопротивляемости,
устойчивости). Если раздражитель продолжает действовать, то может произойти
истощение защитных ресурсов, заканчивающееся болезнью и гибелью. Этот последний
этап многие рассматривают как завершающий период общего адаптационного синдрома
— фазу истощения, или дезадаптации.
В первую «аварийную» фазу для осуществления
компенсаторных реакций развивается мобилизация защитных сил, происходит
вовлечение висцеральных систем разного назначения: кровообращения, дыхания и
симпато—адреналовой системы. Возникают существенные сдвиги метаболизма, носящие
катаболический характер, обеспечивающие организм необходимой энергией, как бы в
предвидении грядущих в скором времени новых функциональных затрат. Происходит
усиление активности липаз и увеличение интенсивности свободно—радикального
окисления липидов клеточных мембран ряда органов. Адаптация в эту стадию
реализуется «с хода», но оказывается несовершенной. Интенсивная активность
систем жизнеобеспечения протекает некоординированно, с элементами хаотичности,
ответные реакции генерализованы, неэкономны и часто превышают потребный для
данных условий уровень. Число измененных показателей в деятельности различных
систем неоправданно велико. Тканевые процессы и, тем более, молекулярные
явления в мембранах клеток в эту стадию направленно не изменяются, т. к. для их
перестройки требуется более значительное время.
Переходный к устойчивой адаптации период характеризуется выраженной
стабилизацией состояния висцеральных систем, формированием межсистемных связей,
способствующих адаптации. В ходе этого периода приспособительные реакции
организма постепенно переключаются на более глубокий тканевой уровень.
Гормональный фон видоизменяется, усиливают свое действие гормоны коры надпочечников
— «гормоны адаптации». По мере проявления анаболической фазы стресс—реакции в
клетках органов, осуществляющих долговременную компенсацию дефектов, происходит
усиленный синтез нуклеиновых кислот и белков, наблюдается их компенсаторная
гипертрофия.
Организм не может длительное время находиться в состоянии
тревоги, и если он выживает, наступает фаза устойчивой адаптации, или
резистентности. Она и является собственно адаптационной реакцией и
характеризуется новым сбалансированным уровнем деятельности функциональных
систем, а также тканевых, клеточных и мембранных структур, перестроившихся
благодаря эндогенным механизмам и активации вспомогательных систем. Основными
особенностями этой фазы являются:
1) мобилизация энергетических
ресурсов;
2) повышенный синтез нуклеиновых
кислот, структурных и ферментативных белков;
3) мобилизация иммунной системы
после предшествовавшей иммунодепрессии;
4) избирательное обеспечение
пластическими и энергетическими материалами тех органов и систем, которые
играют ведущую роль в осуществлении адаптации.
Фаза стойкой адаптации связана с
постоянным напряжением управляющих механизмов, и хотя они более или менее
скоординированы и минимизированы, их бесконечное функционирование невозможно.
Несмотря на экономичность — выключение «лишних» реакций, а, следовательно, и
излишней траты энергии — переключение организма на новый уровень резистентности
не дается даром, а протекает при определенном напряжении, что принято называть
«ценой адаптации».
Формирование устойчивой адаптации
осуществляется при потенцирующем участии стресс—реакции, которая благодаря
действию стресс—гормонов играет важную роль на этапе перехода срочной адаптации
в долговременную. Устойчивая долговременная адаптация к одному фактору нередко
защищает организм от действия многих других факторов. Это явление получило
название «перекрестный защитный эффект адаптации». Например, тренировка к
гипоксии повышает резистентность к большим мышечным нагрузкам и, наоборот,
постоянные мышечные тренировки вызывают эффект устойчивости к гипоксии. На
формирование адаптации большое влияние оказывает и режим воздействия
стрессорного фактора на организм.
Адаптация происходит быстрее, если
стрессор действует не в виде непрерывного фактора, а дискретно. Прерывистый
характер воздействия используется в практике при выработке адаптации к мышечным
нагрузкам, гипоксии, холоду и др.
Общий адаптационный синдром является следствием
функционального напряжения стресс—реализующей системы. Она проявляется
активацией ядер гипоталамуса, нейросекреторные клетки которого усиливают
секрецию либеринов. Они, в свою очередь, стимулируют выработку аденогипофизом
кортикотропина и других тропных гормонов, возрастает поступление в кровь
кортикостероидов и катехоламинов из надпочечников. Эта группа гормонов стимулирует
секрецию тиреоидных и паратиреоидных гормонов, глюкагона, соматотропина,
тирокальцитонина, фермента ренина, угнетая выработку инсулина и половых
гормонов.
Повреждающие факторы активируют также и механизмы стресс—лимитирующих
систем, ограничивающие выраженность стресс—реакции. К наиболее изученным стресс—лимитирующим
системам, препятствующим развитию избыточно выраженной стресс—реакции,
относятся: ГАМК—ергическая, бензадиазепиновая, опиоидергическая,
простагландиновая, серотонинергическая и антиоксидантная.
Известны и некоторые способы коррекции адаптивно—компенсаторных
реакций организма. Их сложный и многокомпонентный характер определяет
многообразие используемых приемов и препаратов. Для профилактики и лечения
повреждающих эффектов стресса обычно используются: препараты общеукрепляющего
действия — адаптогены, витамины, препараты посредников и метаболитов стресс—лимитирующих
систем, препараты вилочковой железы. Для развития антистрессовой устойчивости
доказана высокая эффективность дозированно возрастающих стрессорных факторов,
например, разных модификаций ритмической гипоксической тренировки. В последнее
время широкое распространение получил метод транскраниальной электростимуляции,
вызывающий мобилизацию эндогенных опиоидных пептидов. Можно полагать, что подобного
же эффекта достигали еще в древности, используя приемы акупунктуры. Наконец,
для мобилизации развития общего адаптационного синдрома прибегают к
использованию препаратов кортикотропина, кортикостероидов и др.
ЗАКЛЮЧИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
Организм постоянно приспосабливается к изменяющимся условиям
окружающей среды. Процесс приспособления обеспечивает совокупность
физиологических процессов. В основе их регуляции лежит взаимосвязь всех органов
и функциональных систем организма, что обеспечивает согласованные ответные
реакции.
В процессе эволюции наиболее древней формой взаимодействия
между клетками является гуморальная — посредством веществ, выделяющихся
в жидкости организма. В дальнейшем возникли эндокринные клетки и железы,
которые вырабатывают гормоны, оказывающие влияние на функции организма.
Произошло развитие механизмов нервной регуляции функций, обеспечивающей точную
направленность и быстрое действие, скорость которого значительно выше, чем
действие гормонов.
Взаимодействие нервной и гормональной систем создает единый
интегративный механизм и обеспечивает осуществление всех функций организма, его
приспособление в условиях изменяющейся внешней и внутренней среды. Нервные
клетки способны продуцировать секрет, который содержит гормоны, поступающие во
внутреннюю среду и регулирующие функции организма; это явление называется нейросекрецией.
У беспозвоночных животных вначале преобладающее большинство гормонов
вырабатываются клетками нервных ганглиев; затем развиваются железы эпителиального
происхождения, которые приобретают высокую степень специализации.
У всех позвоночных животных в гипоталамусе существуют
нейросекреторные центры, которые вырабатывают гормоны, оказывающие
влияние непосредственно на органы— или ткани—мишени (антидиуретический гормон,
или вазопрессин, окситоцин и др.) либо действующие на гипофиз и регулирующие
функцию его клеток (гипоталамические рилизинг— и ингибирующие гормоны). Гормоны
аденогипофиза стимулируют действие эндокринных желез, вырабатывающих собственные
гормоны, или оказывают непосредственное влияние на ряд органов и тканей.
Медиаторы тоже являются продуктами нервных клеток и выделяются
окончаниями аксонов в синаптическую щель. Показано, что одни и те же вещества
могут функционировать в качестве медиатора и гормона (например, катехоламины).
Для ряда гормонов в настоящее время установлена множественная локализация
биосинтеза, например ряд гормонов желудочно—кишечного тракта обнаруживается в
мозгу, например, холецистокинин — панкреозимин, бомбезин, субстанция Р,
нейротензин и другие гормоны. Вещества, которые известны у позвоночных как
гормоны, найдены также у многих беспозвоночных животных и у растений.
Предполагают, что в эволюции .рано возникли молекулы, способные выполнять
широкие регуляторные функции; это были ферменты, медиаторы. В дальнейшем часть
из них стала выполнять функции гормонов. Возникновение эндокринных желез,
очевидно, произошло на более поздних этапах эволюции.
В настоящее время у позвоночных животных основная линия
взаимодействия такова: ЦНС → гипоталамус → периферические
эндокринные железы →ткани. Все элементы этой системы объединены обратными
связями, осуществляющимися на разных уровнях. Гипоталамус интегрирует всю
информацию, получаемую от высших отделов мозга и от внутренней среды, и
передает свои влияния эндокринной системе, таким образом наиболее тесно
осуществляется взаимодействие нервной и эндокринной систем.
Основным механизмом поддержания жизнедеятельности организма
на относительно постоянном уровне (гомеостаз) является саморегуляция
физиологических функций, осуществляющаяся на основе единства гуморальных и
нервных механизмов регуляции. У высших животных и человека гомеостаз
поддерживается более строго, чем у низших животных. Это позволило довести до
высокой степени совершенства структуру и функции ЦНС, однако увеличило расход
энергии на гомеостатирование организма. Так, например у млекопитающих и птиц
была создана система терморегуляции, отсутствующая у представителей других
классов и типов, поддерживающая температуру тела с точностью до нескольких
десятых градуса Цельсия. Физиологическая саморегуляция функций является
автоматическим процессом. Значение механизмов обратной связи в поддержании
гомеостаза чрезвычайно велико.
В регуляции функций большое значение имеет также рефлекторный
принцип. Основой осуществления рефлекторных реакций является рефлекторная
дуга, включающая пять основных звеньев. В ряде случаев поступающие в кровь
физиологически активные вещества (медиаторы и гормоны) составляют гуморальное
звено рефлекторной дуги. В регуляции функций большое значение имеет также
феномен доминанты — временно господствующая рефлекторная система, направляющая
работу нервных центров в определенный период.
Ананин
В. Ф. Биорегуляция
человека. В 10 т. М., 1994.
Анохин
П. К. Узловые
вопросы теории функциональной системы. М., 1980.
Бакл
Дж. Гормоны
животных. М., 1986.
Држевецкая
И. А. Основы
физиологии обмена веществ и эндокринной системы. М., 1994.
Дильман
В. М. Большие биологические
часы: Введение в интегративную медицину. М., 1986.
Заварзин А. А., Харазова А. Д.,
Молитвин М. Н.
Биология клетки. Общая цитология. СПб., 1992.
Кассиль
Г. Н. Внутренняя
среда организма. М., 1983.
Новосельцев
В. Н. Теория
управления и биосистемы. СПб., 1994.
Ноздрачев
А. Д. Физиология
вегетативной нервной системы. Л., 1983.
Ноздрачев
А. Д., Чернышева М. П. Висцеральные рефлексы. Л., 1989.
Потемкин
В. В.
Эндокринология. М., 1986.
Розен
В. Б. Основы
эндокринологии. М., 1980.
Селье
Г. Очерки об адаптационном
синдроме. М., I960.
Селье
Г. Стресс без
дистресса. М., 1982.
Телемедицина. Новые информационные
технологии на пороге XXI века / Под ред. Р. М. Юсупова,
Р. И.
Полонникова. СПб., 1998.
Ткачук
В. А. Введение в молекулярную
эндокринологию. М., 1983.
Филаретов
А. А., Подвигина Т. Т., Филаретова Л. П. Адаптация как функция гипофизарно—адренокортикальной
системы. СПб., 1994.
Харди
Р. Гомеостаз. М.,
1986.
Чернышева
М. П. Гормоны
животных: Введение в физиологическую эндокринологию. СПб., 1995.
Юсупов
Р. М., Заболотский
В. П. Научно—методические основы информатизации. СПб., 2000.
Яковлев
Г. М., Новиков В. С., Хавинсон В. X. Резистентность, стресс, регуляция. Л., 1990.