14 РЕПРОДУКТИВНЫЕ ФУНКЦИИ
V—IV вв. до н. э. — Гиппократ (Древняя Греция) в трактате «О семени и
природе ребенка» суммировал знания своего времени о половом акте, зачатии,
беременности. В трактате «О женских болезнях» подробно описал нарушения
менструаций.
IV в. до н. э. —
Аристотель (Древняя Греция) в «Истории животных» дал описание процессов
полового созревания, менструации, полового поведения, зачатия, плодных оболочек
пуповины, положения плода, родов и лактации.
I в. до н. э. —
Древний Рим — начали производить кесарево сечение (от лат. Lex Caesaria —царский, кесарев закон, разрешавший
производить эту манипуляцию) — хирургическую операцию извлечения плода и
последа из полости матки через разрез в ее стенке.
1012—1024 —
Авиценна (Абу Али ибн Сина, Центральная Азия, Иран) в «Каноне врачебной науки»
описал правила здоровой половой жизни, болезней половых органов и способы их
лечения, а также средства увеличения половой потенции, усиления лактации и мн.
др. «Страстную любовь» включил в число «заболеваний, вредящих способности
ощущать и мыслить» и назвал средства лечения этого недуга.
1651 — У. Гарвей
(W. Harvey, Англия) в «Исследовании о
зарождении жизни» предложил термин «эпигенез».
1677 — А. Левенгук (A. Leeuwenhoek, Нидерланды) с помощью микроскопа
открыл сперматозоиды.
сер. XVII в. — Л.
Спалланцани (L. Spallanzani, Италия) осуществил искусственное оплодотворение у
земноводных и млекопитающих.
1825 — Я.
Пуркинье (Пуркине, J. Purkinje, Чехия) открыл яйцо яйцеклетки. 1827 — К. М. Бэр (Россия) подробно
описал яйцеклетку млекопитающих.
1849 — А.
Бертольд (A. Berthold, Германия) пересадил кастрированному петуху семенники
другого петуха и обнаружил исчезновение последствий кастрации.
1858 — К. Экхард
(С. Eckhard, Германия) положил начало научному изучению лактации.
1889 — Ш. Браун—Секар
(Ch. Brown—Sequard, Франция) в опытах на себе показал,
что введение вытяжек из семенников животных оказывает влияние на вторичные
половые признаки.
1894 — И. П.
Павлов (Россия) показал значение нервной регуляции для работы молочной железы.
1905 — 3. Фрейд (Z. Freud, Австро—Венгрия) опубликовал «Три
очерка по теории сексуальности» — первую из многих своих работ по сексологии.
1934 — А.
Бутенандт (A. Butenandt, Германия) с сотр. получили в кристаллическом виде
прогестерон — гормон, подготавливающий матку к имплантации оплодотворенного
яйца.
1935 — А.
Бутенандт и Л. Ружичка (L. Ruzicka, Швейцария) независимо друг от
друга синтезировали мужской половой гормон — тестостерон.
1937 — А.
Мейкпис, Дж. Уэйнстайн и М. Фридман (A. Makepeace, J. Weinstein, M. Friedman, (США) обнаружили, что введение
прогестерона препятствует овуляции у лабораторных животных.
1948—1953 — А.
Кинзи (A. Kinsey, США) опубликовал труды «Сексуальное поведение мужчины» и «Сексуальное
поведение женщины», содержащие анализ опроса 19 тыс. человек.
1955 — группа под
руководством Г. Пинкуса (G. Pinkus, США) создала первый оральный контрацептив.
Под репродуктивными функциями принято понимать комплекс
процессов, который охватывает дифференцировку и созревание половых клеток,
формирование половых мотиваций, половое поведение, половой акт, процесс
оплодотворения, беременность, роды, лактацию, последующее выращивание
потомства. Взаимодействие этих процессов и их регуляция обеспечиваются
системой, центром которой является нейроэндокринный комплекс — гипоталамус —
гипофиз — половые железы (см. разд. 6.2, 6.3.3).
Половые железы
в организме играют двойную роль. Во—первых, в них происходят сложные процессы
сперматогенеза и овогенеза. Итогом этих процессов является развитие к моменту
полового созревания зрелых сперматозоидов и яйцеклеток. Во—вторых, в составе
половых желез находятся особые комплексы с эндокринной функцией. В них происходит
образование половых гормонов, связанных не только с регуляцией функции полового
аппарата; они оказывают влияние на все виды деятельности организма, имеющие
отношение к репродуктивному процессу и соматическому развитию.
Действие
половых гормонов проявляется уже на ранних стадиях эмбрионального развития,
определяя дифференцировку организма в направлении того или иного пола, полового
аппарата и строения ряда систем. Особенно резко влияние гормонов сказывается во
время полового созревания.
Как
известно, у новорожденных пол определяют по строению наружных половых органов.
В процессе полового созревания степень полового диморфизма значительно
возрастает: появляются вторичные половые признаки — лобковое, подмышечное,
лицевое оволосение, мутации голоса, увеличение размеров молочных желез,
формирование специфических пропорций тела, свойственных зрелым мужским и
женским организмам. У животных возникают различного рода выросты типа
петушиного гребня, шпор и пр. С момента полового созревания до угасания
репродуктивной функций половые гормоны определяют цикличность изменений в
организме самок и отчасти самцов. Особенно сложные изменения возникают во время
беременности, развития плода, родов и также поведенческих реакций, связанных с
выращиванием потомства.
Уровнем половых гормонов в крови и их утилизацией в
тканях определяется и функциональная активность гипоталамических центров.
Эта регуляция осуществляется по принципу отрицательных и положительных обратных
связей. В гипоталамусе информация обрабатывается и поступает в его половые
центры. Посредством нейрогормонов (рилизинг—гормонов) гипоталамус
прицельно регулирует секрецию тропных гормонов гипофиза и через нее секрецию
эндокринных желез (см. разд. 6.2). Тем самым в организме обеспечивается
оптимальный для возрастного периода и жизненных условии уровень гормонов.
Половые центры гипоталамуса состоят из тонической и циклической
частей
(центров). У женских индивидуумов под контролем
центров находятся фолликулостимулирующий и лютеинизирующий
гормоны гипофиза, оказывающие регулирующее влияние на половые железы. Тонический
центр поддерживает постоянную базальную секрецию обоих гормонов и
посредством рилизинг—гормонов контролирует синтез и высвобождение
гонадотропинов в количестве, необходимом для развития фолликулов в яичниках
между овуляциями. Циклический центр включается в контроль половых желез
импульсно только для осуществления
овуляции. Он обеспечивает овуляторный выброс лютеинизирующего гормона из
гипофиза, в результате чего происходит разрыв созревших фолликулов.
Тонический центр локализован в медиальном отделе
гипоталамуса, циклический — в предоптической области. Обладая определенной
функциональной автономностью, гипоталамические половые центры вместе с тем
находятся под контролем миндалевидного тела и гиппокампа, нейроны которых
осуществляют рецепцию уровня стероидных гормонов.
Гипоталамические центры являются не только главным
звеном системы регулирующей выработку гормонов, они играют и ведущую роль в
формировании полового поведения. Например, при введении в предоптическую
область кастрированных самцов животных тестостерона — мужского полового
гормона. обладающего наибольшей андрогенной активностью, у них происходит
восстановление всего комплекса поведенческих реакций, связанных с копуляцией.
Тестостерону свойственно много функций: осуществление обратной связи с
гипоталамусом и передней долей гипофиза, обеспечение сперматогенеза, регуляция
сексуального поведения, формирование вторичных половых признаков. Семенники
продуцируют также относящийся к классу гликопротеинов гормон ингибин, с
помощью которого регулируется высвобождение в передней доле гипофиза
фолликулостимулирующего гормона.
У самок разрушение предоптических и передних
гипоталамических ядер резко угнетает половую активность и сопровождается полной
утратой способности к спариванию. Напротив, раздражение латерального
гипоталамического поля сильно стимулирует половое поведение у самцов.
Раздражение задних отделов вентромедиального гипоталамического ядра, связанного
с передней гипоталамической областью, резко повышает половое влечение и
вызывает преждевременную овуляцию.
Уровнем половых гормонов регулируется секреция
гонадотропин—рилизинг—гормона (ГнРГ). Нейроны, в которых образуется ГнРГ,
распределены в нескольких зонах гипоталамуса. Поступающая из внешней и
внутренней среды организма информация передается в гипоталамус, где
синтезируется гуморальный мессенджер ГнРГ, являющийся по структуре
декапептидом. Нейроны продуцирующие ГнРГ, развиваются из обонятельной пластинки
и в процессе эмбриогенеза мигрируют и рассеиваются в предоптической зоне,
перегородке диагональной полоске (Брока), но наибольшая их концентрация
определяется в медиобазальной зоне гипоталамуса, в области воронки и
дугообразного ядра.
Эндокринная функция половых желез выражена не
одинаково у животных разных типов. У беспозвоночных она развита, по—видимому,
слабо, хотя появление у кольчатых червей и ракообразных вторичных половых
признаков коррелирует с выработкой половой железой биологически активных
веществ
У рыб, амфибий и рептилий эндокринная функция половых
желез а также связанные с ней вторичные половые признаки и инстинкты получают
отчетливое выражение. Кастрация вызывает у них недоразвитие или обратное
развитие вторичных половых признаков и исчезновение связанных с размножением
инстинктивных реакций. Особенно яркое выражение такой зависимости проявляется у
птиц. Так, ранняя кастрация петухов и кур приводит к развитию организма по
бесполому типу, в результате самцы и самки внешне становятся почти нeoтличимы.
У петуха исчезает
типичная постановка корпуса, атрофируется гребень, пропадают половые инстинкты.
У курицы оперение становится петушиным, отрастают шпоры, гребень приобретает
вид гребня кастрированного петуха.
Значительное усложнение эндокринной функции, особенно
регулирующей половую деятельность самки, является характерным признаком
млекопитающих. Как известно, рыбы, амфибии, рептилии, птицы являются
яйцекладущими У млекопитающих возникла новая биологическая особенность —
развитие зародыша внутри материнского организма и живорождение. Все эти
процессы обеспечиваются новыми механизмами, связанными в первую очередь с
эндокринными железами. Среди них определенное значение принадлежит периодически
возникающему в яичниках млекопитающих желтому телу и гормонам плаценты (см.
разд. 6.3.3). У амфибий, рептилий, птиц настоящих желтых тел
еще нет.
В процессе эволюционного
развития у позвоночных животных половые железы, или гонады, развиваются у края
первичной почки. Половые клетки выходят через протоки, образующиеся из каналов
выделительной системы — мезонефрального и парамезонефрального. У мужских особей
высших позвоночных из мезонефрального протока развиваются пути выведения семени
— проток придатка, семявыносящий и семявыбрасывающий протоки. У особей женского
пола мезонефральный проток редуцируется, а из парамезонефрального образуются
маточные трубы, матка, влагалище.
У млекопитающих половые железы — яичники у
женских особей, семенники у мужских — органы овальной или бобовидной
формы. Яичники располагаются в полости таза, семенники у большинства
млекопитающих перемещаются в особое образование брюшной полости— мошонку.
Мочеполовая пазуха высших млекопитающих животных открывается самостоятельно
наружу и образует у особей женского пола преддверие влагалища, мужского пола — мочеполовой канал.
Характерной особенностью млекопитающих является
срастание дистальных концов яйцеводов и образование у плацентарных животных
непарного влагалища, а у большинства также и матки. У некоторых насекомоядных,
грызунов и слонов матка «двойная», хищных — «двураздельная», у большинства
млекопитающих — «двурогая». Окончательное срастание дистальных концов яйцеводов
и образование матки с одной полостью, открывающейся во влагалище, произошло у
ряда рукокрылых, обезьян и человека. Парность у них сохранили только начальные
отделы яйцеводов — маточные трубы.
У млекопитающих происходит образование добавочных
желез. Крупные пузырчатые железы — семенные пузырьки — располагаются в конце
семявыносящего канала; предстательная железа, бульбоуретральные железы — в стенках
мочеполового канала самцов. Большие железы преддверия влагалища (бартолиновы
железы) формируются у особей женского пола.
У мужчин половой железой является яичко (тестис) и
придаток яичка. Этот парный орган расположен в мошонке. От протока придатка
начинается семявыносящий проток. Он проходит в составе семенного
канатика через паховый канал, а затем опускается по стенке малого таза. В малом
тазу расположены семенные пузырьки. Их выделительные протоки соединяются
с семявыносящими протоками и образуют правый и левый семявыбрасывающие
протоки. Последние открываются в мочеиспускательный канал, и таким образом
здесь мочевой путь соединяется с половым. В месте впадения протоки прободают
непарную предстательную железу, охватывающую мочеиспускательный канал.
Мочеиспускательный канал далее проходит в губчатом теле полового члена, где в
него впадают протоки бульбоуретральных желез, и оканчивается наружным
отверстием.
|
|
|
Упрощенная схема строения мужских половых органов 1 — яичко, 2 —
придаток яичка, 3 — пещеристая ткань, 4
— головка полового члена, 5 — лобковый симфиз, 6 — мочевой пузырь, 7
— семявыносящий проток, 8 —
предстательная железа, 9 —
семенной пузырек. |
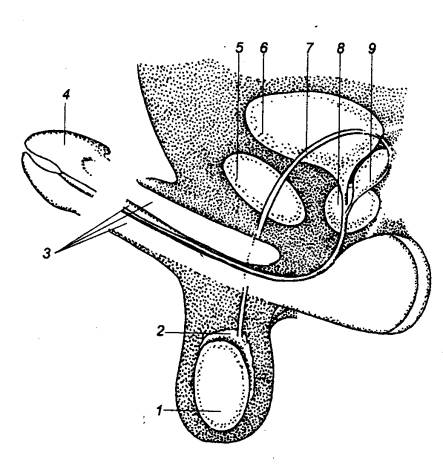
Перед рождением яички опускаются из брюшной полости через
паховый канал в мошонку, что имеет исключительно важное значение для создания
температурных условий на 2—3 °С ниже температуры ядра тела. Указанная разница в
температуре необходима для сперматогенеза. Осуществлению этой разницы
способствует два механизма. Во—первых, это венозное сплетение которое служит в
качестве противоточного теплообменника между теплой артериальной кровью,
поступающей к яичкам, и более холодной, оттекающей от них венозной кровью. Во—вторых,
в зависимости от состояния мышцы, поднимающей яичко (сокращенная или
расслабленная) регулируется оптимальное расстояние в положении яичка по
отношению к туловищу. Если яички находятся продолжительное время при повышенной
температуре, например, при нарушении терморегуляции или при лихорадке, то это
может привести к временному или постоянному бесплодию в результате нарушения
сперматогенеза.
Из позвоночных копулятивный орган
имеют животные, для которых характерно внутреннее оплодотворение и живорождение.
У живородящей собачьей акулы в копулятивный орган преобразовались внутренние
части брюшных плавников, у костистых рыб — анальный плавник. У змей и ящериц
копулятивным органом являются выпячивания кожи у края клоаки. У крокодилов и
черепах имеется непарное утолщение стенки клоаки, образованное пещеристой
тканью. Этот копулятивный орган снабжен желобком для отекания семени.
Аналогичный по строению орган существует у большинства видов птиц.
Млекопитающие характеризуются разным
строением копулятивного органа. Так, у яйцекладущих половой член находится
внутри клоаки, он выдвигается и втягивается специальными мышцами, бороздка на
нем превратилась в канал. Половой член у сумчатых располагается вблизи заднего
прохода, окружен общим с ним сфинктером, направлен назад. Конец его раздвоен
что соответствует двойному влагалищу самки. У других млекопитающих половой член
направлен вперед. У сумчатых, ластоногих, китообразных, рукокрылых, отдельных
хищников и грызунов, полуобезьян и некоторых обезьян помимо кавернозных и
губчатых тел в передней части органа лежит кость полового члена, а у некоторых
животных находятся ароматические железы.
У человека в половом члене
различают тело, корень и головку (рис. 14.1). Тело состоит из двух
пещеристых тел и губчатого тела, заключающего часть мочеиспускательного канала.
Проксимальные концы пещеристых тел — ножки полового члена — закрепляются на
лобковых и седалищных костях, дистальные концы заострены и входят под венец
головки. Губчатое тело прилежит к нижней поверхности спаренных пещеристых тел.
Его задний конец уплощен и носит название луковицы полового члена, а
передний переходит в головку полового члена. Головка имеет вид
конусообразной изогнутой пластинки. На заостренной верхушке головки открывается
наружное отверстие мочеиспускательного канала.
|
|
|
Рис. 14.2 Упрощенная схема строения женской половой
системы 1 — маточная труба, 2 — полость матки, 3 — шейка матки,
4 — влагалище, 5 — клитор, 6 — отверстие мочеиспускательного канала,
7 — задний проход, 8 —
отверстие влагалища, 9 —
малая губа, 10 — большая
губа, 11 — наружные половые
органы (вульва), 12 — тело
матки, 13 — складка брюшины, 14 — правый яичник, 15 — отверстие маточной трубы, 16 — бахромка. |
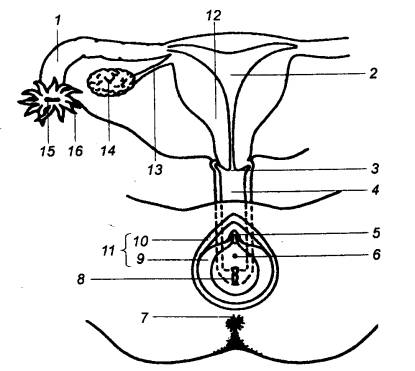
Окружающая головку кожа образует
складку, называемую крайней плотью. Ее вентральный листок получил
название уздечки крайней плоти. Источниками кровоснабжения полового
члена являются ветви внутренней половой (срамной) и уретральной артерий. Все они соединены между собой
анастомозами. Соматическая иннервация обеспечивается ветвями полового
(срамного) нерва и тыльным нервом полового члена. Симпатическая иннервация
происходит в основном из нижнего брыжеечного сплетения, парасимпатическая — из
тазового нерва.
У женщин половой железой являются яичники (рис.
14.2). Они расположены в полости малого таза у боковых стенок. Средние размеры
яичников у женщин зрелого возраста таковы: длина — 3—4 см, ширина — 2—2,5,
толщина — 1—1,5 см, масса — 6—8 г. В яичнике различают маточный и трубный концы
Трубный конец приподнят кверху и обращен к воронке маточной
(фаллопиевой) трубы. Яичник подвижно соединен связками с маткой и
стенкой таза.
Яичники низших беспозвоночных представляют лишь
временное скопление половых клеток. У более высоко организованных животных они
становятся обособленными органами. У низших червей, иглокожих, членистоногих,
моллюсков яичники мешкообразные. Яйца в них образуются во внутренней части
органа и после созревания по специальным каналам выводятся наружу.
Способность к оплодотворению у женщин, в отличие от
мужчин, изменяется циклически, так как образование зрелых яйцеклеток происходит
у них нерегулярно. В противоположность мужскому организму, в котором ежедневно
образуются миллионы гамет, в женском созревает только одна или несколько
яйцеклеток, и то в определенное время.
У всех позвоночных созревшие яйца через прорыв стенки
яичника попадают в общую (вторичную) полость тела и оттуда выводятся через
яйцеводы. Фолликулы в яичнике позвоночных содержат одну крупную яйцеклетку,
окруженную мелкими фолликулярными клетками, которые служат для питания
яйцеклетки и образования яйцевой оболочки. У низших позвоночных образование
яйцеклеток происходит в течение всей жизни, у высших — только в эмбриональном
периоде или сразу после рождения. Внутри фолликула у млекопитающих образуется
полость, наполненная серозной жидкостью. В момент созревания яйцеклетки
фолликул лопается и яйцеклетка попадает в полость тела, на месте же фолликула
образуется эндокринная
структура — желтое тело.
Как и мужские гонады, яичники
выполняют двойную функцию: образование половых клеток (овогенез) и синтез
женских половых гормонов (стероидогенез). Основными половыми гормонами у женщин
являются эстрадиол, который образуется при развитии фолликула, и прогестерон,
синтезируемый в желтом теле. Гормоны различаются по структуре и выполняемым
функциям, однако их эффекты
могут быть и синергичными, и антагонистичными. В яичниках также образуется
полипептидный гормон ингибин— регулирующий секрецию
фолликулостимулирующего гормона.
Оплодотворение
яйцеклетки обычно происходит в маточной (фаллопиевой) трубе — парном трубчатом
органе, осуществляющем функции транспортировки яйцеклетки и сперматозоидов,
создания благоприятных условий для оплодотворения, развития яйцеклетки в ранние
сроки беременности и продвижения зародыша первых дней развития в матку.
Маточная труба одним концом открывается в матку, другим — в полость брюшины
вблизи яичников. Брюшное отверстие, диаметр которого 2—3 мм, обычно
сомкнуто. Его открытие связано с процессом овуляции. Во время овуляции брюшной
конец маточной трубы может плотно соприкасаться с яичником. В маточной трубе выделяют воронку,
ампулу и перешеек. Воронка открывается в полость брюшины, ее ворсинки
захватывают яйцеклетку во время овуляции и далее способствуют продвижению в
ампулу. Ампула именно то место где происходит оплодотворение. Она имеет
слабо выраженный мышечный слой и сильно развитый эпителий. Перешеек
расположен в области соединения трубы и матки и представляет собой полый
просвет, являющийся механическим препятствием для движения клеток.
В маточных трубах осуществляется
транспорт половых клеток во встречных направлениях. Сперматозоиды движутся из
матки к ампуле, а возникшие после оплодотворения зиготы — в маточную полость.
Координация сокращений гладкой мускулатуры и степень движений ресничек требует
тонкой координации что достигается посредством специальных гормональных и
нервных влияний.
Матка имеет
грушевидную форму, обращена узким концом в верхний отдел влагалища. В матке
различают дно, тело, шейку и полость. Дном называется верхняя часть
матки выше отхождения маточных труб. Тело имеет треугольную форму, его
продолжением, составляющим нижнюю часть, является шейка матки. Полость
матки рожавшей женщины на фронтальном разрезе имеет треугольную форму. В
верхних углах этого треугольника находятся отверстия, открывающиеся в маточные
трубы, в нижнем углу — перешеек ведущий в полость канала шейки матки.
Шейка матки конической или цилиндрической
формы. В нижнем ее конце канал открывается во влагалище.
Влагалище —
мышечно—эластическая трубка, расположенная в малом тазу верхним концом она
охватывает шейку матки, нижним — заканчивается в области преддверия влагалища.
У девственниц дно преддверия и его нижний конец ограничены девственной
плевой. Направляясь из полости таза к преддверию, влагалище проходит через
мочеполовую диафрагму. Влагалище участвует в процессах совокупления и
оплодотворения, в родах составляет часть родового канала. Длина влагалища у
зрелой женщины колеблется от 7 до 9 см ширина — 2—3 см, задняя стенка на 1,5—2
см длиннее передней. Влагалище может менять свою форму, диаметр и глубину при
сокращении мышц тазового дна, матки и мышечных элементов связочного аппарата.
Функционально влагалище делится на две части: верхнюю
и нижнюю верхняя часть расширена, она способна к активному сокращению нижняя
сужена и более массивна.
В период
полового возбуждения происходит резкое кровенаполнение вен влагалища, удлинение
его верхних частей, увеличение транссудации в просвет влагалища. После полового
акта слизистая оболочка влагалища способна всасывать плазму спермы и
вырабатываемые семенными пузырьками простагландины. Во время родов влагалище
сильно растягивается, но спустя неделю после них благодаря эластичности стенок
происходит сокращение влагалища, хотя
просвет его остается более широким, чем до родов.
Книзу от
мочеполовой диафрагмы, замыкающей выход из малого таза. расположены наружные
женские половые органы. Они включают женскую половую область
(вульву). К женской половой области относятся лобок, большие и малые половые
губы, клитор, преддверие влагалища, его железы, луковица преддверия. Разделение
половых органов на наружные и внутренние обьясняется не только особенностями их
топографии, но и спецификой эмбрионального развития и функции. Развитие женских
половых органов происходит частично за счет кожных покровов нижнего отдела
туловища.
Лобок является самым нижним участком брюшной стенки. Он
имеет форму треугольника, основание которого направлено книзу. Лобок переходит
в большие половые губы. Большие половые губы представляют парные
парасагиттально расположенные валики кожи, в толще которой заложена жировая
клетчатка с заключенным в ней венозным сплетением и пучками эластических
волокон. Малые половые губы располагаются кнутри от больших и
параллельно им. В их толще также имеется соединительная ткань и относительно
крупное венозное сплетение. Вместе с большими половыми губами они ограничивают
с боков половую щель. В переднем углу половой щели между малыми половыми
губами находится клитор, в толще которого лежит пещеристое тело.
Несколько кзади от клитора, между ним и входом во влагалище, расположено наружное
отверстие мочеиспускательного канала, которое открывается в преддверие
влагалища. Дно преддверия образовано девственной плевой. Основу девственной
плевы составляет соединительная ткань с эластическими, коллагеновыми и
мышечными волокнами, создающими ее тургор. В основании и толще больших половых
губ помещаются две доли непарного пещеристого образования — луковицы
преддверия.
В функциональном отношении женская половая область
является по преимуществу органом полового чувства. В клиторе находится большое
число механорецепторов. Во время полового возбуждения клитор набухает. Это
происходит вследствие усиления притока артериальной крови и ослабления оттока
венозной. Параллельно с этим набухает и луковица преддверия, представляющая
собой венозное сплетение, напоминающее кавернозное тело. В этот момент из желез
преддверия выделяется богатый муцином секрет, увлажняющий вход во влагалище.
Половое созревание представляет процесс формирования
репродуктивной функции женского и мужского организмов, которое завершается половой
зрелостью — способностью к воспроизводству полноценного потомства.
В половом созревании принято
различать три фазы: препубертатную, собственно пубертатную и
постпубертатную. Первая охватывает два—три года, предшествующие появлению
признаков полового созревания. Вторая протекает от начала лобкового оволосения,
увеличения яичек у мальчиков, набухания молочных желез и их легкой
болезненности у девочек до первых поллюций у мальчиков и первой менструации у
девочек, третья — до появления так называемых юношеских угрей и начала оволосения
лица у мальчиков.
С периода полового созревания
начинается процесс спермато— и овогенеза. Он продолжается всю жизнь до самой
старости. Мужские и женские половые клетки различаются онтогенетически,
количественно и морфологически. Сперматозоиды имеют много общих свойств с
овоцитами, включая общее происхождение, взаимодействие с окружающими клетками
(клетки Сертоли и фолликулярные клетки), частичный контроль со стороны
гонадотропных гормонов и зависимость от половых стероидных гормонов. Обе клетки
образуются в результате мейоза и имеют гаплоидный набор хромосом. Однако между
этими клетками имеются и существенные различия в сроках образования в процессе
онтогенеза, количестве образующихся гамет и морфологии.
В сперматогенезе
принято рассматривать три стадии — митоз, мейоз и дифференцировка клеток
(спермиогенез). У мужчин процесс сперматогенеза продолжается 65—70 дней.
Происходит он на протяжении всех семенных канальцев. Новый цикл начинается
через одинаковые временные интервалы поэтому вдоль каждого канальца можно
увидеть клетки на разных стадиях развития. Именно таким образом поддерживается
продолжительная непрерывная продукция сперматозоидов. Каждые сутки их
образуется около 2 х 108 Сперматогонии в мужском организме
продолжают делиться с начала полового созревания до старости. У женщин в
течение жизни образуется лишь несколько сотен зрелых яйцеклеток, и процесс этот
прекращается в период менопаузы.
Сперматозоиды — клетки небольшого размера, диаметр
их составляет 1—2 мкм. Форма их хорошо приспособлена для перемещения и
взаимодействия с яйцеклеткой. В результате мейоза из каждого сперматогония
образуется четыре одинаковых сперматозоида. В головке сперматозоида находится
ядро, содержащее гаплоидное число хромосом. Оно прикрыто акросомой,
представляющей собой особую ограниченную мебраной структуру, содержащую
гидролитические ферменты. Ферменты способствуют проникновению сперматозоида в
яйцеклетку непосредственно перед оплодотворением. Функционально ее иногда
рассматривают как увеличенную лизосому.
Яйцеклетка имеет размер около 100 мкм в
диаметре. Она является самой крупной клеткой организма и внешне не проявляет
признаков специализации. Каждый овогоний дает начало только одной яйцеклетке.
В период полового
созревания происходит формирование вторичных половых признаков и фенотипа.
Непосредственное участие в этом принимают андрогены. Они определяют
распределение волос, особенности кожи, рост костей развитие мышц. Андрогены у
мужчин стимулируют рост волос на лице, груди и в подмышечных впадинах. Однако в
сочетании с генетическим фактором обеспечивается формирование залысин в
височных областях. Рост волос в подмышечных впадинах и на лобке у женщин также
определяется андрогенами. Увеличение продукции андрогенов у женщин приводит к гирсутизму
— избыточному росту волос по мужскому типу.
Возрастание чувствительности клеток—мишеней
к андрогенам приводит к развитию юношеских угрей у лиц мужского и женского
пола. Рост гортани и утолщение голосовых связок также зависит от андрогенов,
поэтому евнухи имеют высокий голос, как у мальчиков до начала полового
созревания. Под влиянием андрогенов происходит закрытие эпифизов костей, что
предотвращает дальнейший рост индивида. Поэтому преждевременное половое
созревание, как правило, сочетается с низким ростом, в то время как у людей с
задержкой полового развития и у евнухов рост обычно высокий.
Заключительным этапом полового
созревания организма, характеризующимся общим физическим развитием и таким же
морфофункциональным состоянием половых органов, которые позволяют без ущерба
для здоровья осуществлять половые функции, является половая зрелость.
Срок ее достижения индивидуален. У лиц женского пола он наступает в возрасте 16—18
лет муже кого пола — 18—20 лет.
Через восходящие активирующие
влияния гипоталамических центров половые гормоны направленно воздействуют на
различные структуры головного мозга (включая кору больших полушарий),
ответственные за половое поведение. Половые гормоны тем самым организуют и половое
влечение (либидо) — одну из основных биологических мотиваций, от
удовлетворения которой зависит существование индивида и рода.
В половом влечении различают нейрогуморальный
компонент и половую доминанту. Первый связан с функционированием
врожденных комплексов рефлексов, вторая представляет собой системный феномен.
Она включает прочные связи на основе запечатлевания, условнорефлекторные связи
и психоэмоциональный настрой (см. разд. 3.19.1, 3.26.3). В зависимости от
возрастного периода половое влечение усложняется и может проявляться в
различных формах: понятийной, платонической, эротической, собственно
сексуальной, в форме зрелой сексуальности.
Понятийная форма включает элементарное представление о
половых различиях, например у детей младшего возраста, дифференцирующих пол по таким
условным внешним признакам, как одежда. Платоническая форма указывает на
возникновение высших психологических установок, свободных от интереса к
генитальной сфере. Она проявляется в романтических фантазиях. Эротическая
форма характеризуется стремлением к интимному общению. Едва намеченная у
большинства юношей, у девушек она нередко завершает половое развитие. Собственно
сексуальная форма протекает на фоне специфически окрашенных эмоций с четким
проявлением интереса к генитальной сфере.
Половое влечение как доминирующая
мотивация вызывает в организме значительные сдвиги. Например, возрастает
острота обоняния, зрения, слуха, повышается чувствительность рецепторов кожи и
особенно определенных частей тела — эрогенных зон, раздражение которых
вызывает половое возбуждение. Такими эрогенными зонами у мужчин являются
наружные половые органы. У женщин эрогенной зоной может быть любая часть тела,
но чаще это генитальная область.
Степень выраженности полового влечения
зависит от индивидуальных особенностей организма, личности человека и условий
воспитания, стимулирующих или подавляющих развитие влечения.
Как известно, внутреннее
оплодотворение составляет важную часть репродуктивного цикла у высших животных.
У многих из них, в том числе и человека, оно облегчается благодаря
существованию совокупительного органа — мужского полового члена, который
вводится во влагалище и вносит гаметы как можно глубже в половые пути самки.
Половой акт (коитус, половое сношение,
совокупление, соитие) — физический процесс, начинающийся с момента введения во
влагалище полового члена и заканчивающийся эякуляцией и оргазмом.
Половому акту предшествует нейрогуморальная готовность к нему организма. Эта
готовность складывается во время полового созревания и исчезает с угасанием
репродуктивных функций.
У мужчин в половом акте
различается несколько стадий. Считают что первая из них начинается с постановки
цели полового сближения и выражается в активных действиях, ведущих к интимной
близости. Если обстановка благоприятствует, то возникает следующая — эрекционная
стадия, во время которой наряду с эрекцией полового члена происходит
выделение бульбоуретральными (куперовыми) железами нескольких капель жидкого секрета.
Секрет смачивает головку полового члена, что значительно облегчает его
последующее введение во влагалище. Пусковую роль для возникновения эрекции у
мужчин играет эмоциональное влечение к женщине. Отдельно взятые обонятельные,
зрительные, слуховые, тактильные раздражители сами по себе не всегда способны
вызвать эрекцию. Однако их суммация, постепенно возрастая, достигает порога
возбудимости эрекционного центра, что сопровождается эрекцией.
Третья стадия — фрикционная —
начинается с момента введения полового члена во влагалище. Она характеризуется
последовательным рядом фрикций — возвратно—поступательных движений
полового члена, в результате которых происходит суммация возбуждения,
возникающего в эрогенных зонах. Трение, создаваемое ритмическими движениями,
производит тактильное раздражение в головке полового члена отростков
чувствительных клеток. Это приводит к активации симпатических нейронов, которые
вызывают сокращение внутреннего сфинктера мочевого пузыря, гладкой мускулатуры
придатков яичка, семявыносящего протока и придаточных желез (семенного
пузырька, предстательной железы и бульбоуретральных желез). В результате
происходящего будущие компоненты семенной жидкости выталкиваются в
проксимальную часть уретры. Здесь все они смешиваются. Возросшее давление в
проксимальной части уретры сопровождается возникновением рефлекторной
активности двигательных нейронов, иннервирующих мышцы основания полового члена.
Ритмические волнообразные сокращения этих мышц продвигают секреты желез ближе к
дистальной части уретры.
С достижением порога эякуляции
наступает четвертая — эякуляторная — стадия. Во время этой стадии в
результате мышечных сокращений семявыносящих путей происходит извержение
семени. Процесс семяизвержения возникает на фоне максимального полового возбуждения
и сопровождается сладострастным ощущением — оргазмом. Интенсивность
оргазма зависит от темперамента и конституции человека.
Во время полового акта смазку обеспечивают главным образом
секреты влагалищных желез, частично же в этом процессе участвует и прозрачная
жидкость бульбоуретральных желез мужчин. Секреты придаточных мужских желез
характеризуются щелочной реакцией. Они содержат фруктозу, витамины, лимонную
кислоту, простагландины и широкое разнообразие различных ферментов. Все они
вместе со слизью повышают рН среды влагалища до 6—6,5, что является оптимальным для подвижности
сперматозоидов после эякуляции.
После эякуляции и оргазма размеры полового члена
уменьшаются, наступают периоды абсолютной и относительной рефрактерности. Во
время абсолютной рефрактерности мужчина оказывается нечувствительным к
эротическим стимулам. Во время относительной рефрактерности эрекция может
возникнуть лишь при действии необычайных по силе раздражителей.
|
|
|
Рис. 14.3 Фазы. полового акта мужчины (А) и женщины (Б) I — возбуждение, II
— плато. III — оргазм, IV — спад; 1—5 —
индивидуальные проявления половой активности. |
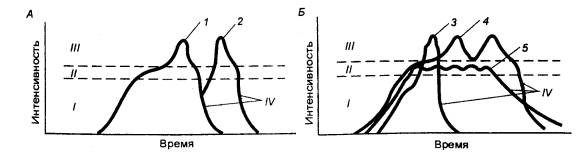
У женщин
сексуальное реагирование выражается в приливе крови к половым органам,
увеличении поперечника малых половых губ, удлинении и расширении влагалища, в
выделении секрета железами его стенки. Выраженность этих признаков указывает на
степень готовности к половому акту. Суммация возбуждений, возникающих при
раздражении эрогенных зон, а также тактильных раздражении, вызываемых фрикциями
полового члена во влагалище, приводит к сужению влагалищного канала и
образованию так называемой оргастической манжетки (см. разд. 14.8.2). Процесс
оргазма нарастает у женщин медленнее, чем у мужчин, и угасание его также
продолжительнее. После оргазма уменьшение набухания половых губ и клитора
происходит быстро, влагалище приобретает обычные размеры.
В половом акте принято
различать ряд фаз — возбуждение, плато, оргазм и спад (рис. 14.3).
У мужчин эти фазы по длительности и интенсивности проходят стереотипно, у
женщин они более разнообразны. Временные соотношения этих фаз сугубо
индивидуальны, однако наиболее продолжительными из них являются фаза
возбуждения и спада, короткими — плато и оргазма.
Понятие половая жизнь
включает сумму соматических, висцеральных, психических, социальных процессов и
отношений, в основе которых заложено и посредством которых удовлетворяется половое
влечение. У человека инстинктивная безусловнорефлекторная основа не имеет
абсолютного определяющего значения, она приобретает качественно новые аспекты и
служит не только цели продолжения рода, но предусматривает личностное
обьединение между мужчиной и женщиной, духовное общение между ними. В развитом
человеческом обществе взаимоотношение полов регулируется системой норм морали.
Формой половой жизни является половое сношение между мужчиной и женщиной,
которое вместе с их духовным единством входит в понятие полноценная половая
жизнь.
Возраст, при котором начинается половая жизнь, определяется половым созреванием.
Частота половых сношений зависит от особенностей личности и от ряда
внешних, главным образом социальных, факторов. При этом особое значение имеет
вступление в брак, после которого половая жизнь, как правило, становится
регулярной. При доступности половых сношений, например в браке, в период зрелой
сексуальности устанавливается определенная ритмичность половой жизни.
Частота половых актов в норме зависит от степени половой
возбудимости, силы полового влечения, выраженности задерживающих влияний и у
разных людей проявляется индивидуально — от единичных до нескольких десятков
половых сношений в неделю. Продолжительность каждого из актов также
вариабельна. У здоровых людей она колеблется в пределах 1,5—5 мин.
Продолжительность половых сношений уменьшается с увеличением интервалов между
ними. Вместе с тем универсальной нормы половой жизни (ее интенсивности
продолжительности) не существует. Она меняется в зависимости от возраста, пола,
личностных особенностей и других факторов. Благополучие половой жизни во
многом зависит от того, насколько гармонично сочетаются индивидуальные
особенности мужчины и женщины, т. е. от наличия психологической и
психосексуальной совместимости партнеров.
В первые месяцы брака половая жизнь, как правило,
интенсивна. Но у этого периода есть своя особенность: чем моложе мужчина, чем
сильнее его половая конституция и чем желаннее для него близость, тем быстрее
наступает оргазм, тем короче половой акт. Иногда это становится предметом
тревоги мужчины. Как считают сексологи, для тревог причин нет, именно в этот
период у молодых женщин преобладают платоническое и эротическое влечения (см.
разд. 14.5). Пока не проснется зрелая сексуальность, красивая прелюдия полового
акта для них не менее, а чаще более приятна, чем сам акт, а продолжительность
его не имеет значения.
Гармония интимной жизни создается не сразу, необходимо
время, чтобы научиться чувствовать друг друга, преодолеть психологические
барьеры. Основным признаком сексуальной зрелости мужчины является переход от
ежедневных повторных половых актов на ритм 1—3 акта в неделю. Устанавливается и
их нормальная продолжительность— от 1,5 до 5 мин. Ритм 1—3 акта в неделю
наиболее естествен, он носит название условнофизиологического.
Установившись в первые месяцы супружества, этот ритм может сохраняться до
старости. В возрасте после 60—65 лет иногда достаточно прекращения интимной
близости на 1 мес, чтобы половая функция не восстановилась. Вместе с тем сохранение
ее крайне желательно, так как способствует повышению общего тонуса, дает
ощущение полноты жизни.
14.8. РЕФЛЕКТОРНЫЕ МЕХАНИЗМЫ ПОЛОВОЙ ДЕЯТЕЛЬНОСТИ
Концепция механизмов половой
деятельности опирается на общую теорию функциональных систем П. К. Анохина —
учения о конечном полезном результате как организующей силе любой
физиологической активности. В основе половой деятельности млекопитающих и
человека лежат комплексы рефлекторных актов. Эти сложные реакции осуществляются
афферентными процессами соматической и висцеральной природы, соматическими,
симпатическими, парасимпатическими и отчасти метасимпатическими эфферентными
воздействиями.
14.8.1. Половые рефлексы у мужчин
Половой цикл у мужчин (табл. 14.1) включает несколько
последовательных фаз: эрекции полового члена, эмиссии спермы и секретов
добавочных половых желез (семенной жидкости) в дистальную часть
мочеиспускательного канала, эякуляции этой жидкости из уретры, оргазма.
Эрекция. Эрекция — увеличение полового члена в объеме и приобретение им
механической твердости, обеспечивающей возможность полового сношения. В основе
эрекции лежит быстрое и длительно сохраняющееся усиление притока артериальной
крови в пещеристые тела полового члена и губчатое тело уретры в сочетании с
замедлением венозного оттока. Преобладание артериального притока над венозным
оттоком возникает в результате существования в просвете артерий полового члена
особых замыкательных валиков.
Расширение артерий обеспечивается сосудодвигательными
парасимпатическими волокнами нейронов крестцового отдела спинного мозга,
проходящими в составе тазового нерва (рис. 14.4). Наступление эрекции
определяется функцией супраспинальных (высших корковых и подкорковых)
механизмов. Возбуждение парасимпатических нейронов поддерживается афферентной
импульсацией с основной эрогенной зоны — головки полового члена. Возбуждение
механорецепторов головки возникает при скользящих движениях — фрикциях во время
полового акта.
|
|
|
Рис. 14.4 Иннервация половых органов мужчины 1 — яичко, 2 —
придаток яичка, 3 — пещеристая ткань, 4
— головка полового члена, 5 — симфиз, 6 — мочевой пузырь, 7
— семявыносящий проток, 8 —
предстательная железа, 9 —
семенной пузырек, 10 — симпатические
нейроны, 11 — грудные и
поясничные сегменты спинного мозга, 12
— парасимпатические нейроны, 13
— крестцовые сегменты спинного мозга, 14 — половой нерв, 15
— соматические эфференты, 16
— внутренностный тазовый нерв; широкой стрелкой обозначены нисходящие
влияния; спинномозговые интернейроны и функциональные модули метасимпатической
нервной системы не показаны. |
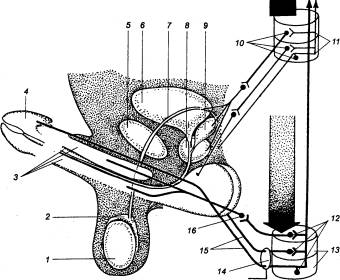
Таблица
14.1
Половой цикл у мужчин
|
|
Эрекция |
Эмиссия и
эякуляция |
Оргазм |
|
Афференты |
От
головки полового члена и окружающих тканей. к крестцовому отделу спинного
мозга (в составе половых нервов) |
От
наружных и внутренних половых органов к крестцовому (в составе половых и
тазовых нервов и пояснично—грудному (из подчревного сплетения) отделам
спинного мозга; афферентные волокна от скелетных мышц |
Возможен,
если сохранен хотя бы один путь поступления чувствительных сигналов (от
половых органов к крестцовому и пояснично—грудному отделам; от скелетных мышц
к крестцовому отделу) |
|
Вегетативные
эфференты |
1)
Парасимпатические из крестцового отдела (рефлекторные и психогенные влияния);
2) симпатические из пояснично—грудного отдела (психогенные влияния) |
Симпатические
из пояснично—грудного отдела |
|
|
Соматические
эфференты |
— |
К
луковично—губчатым и седалищно—пещеристым мышцам и мышцам тазового дна |
|
|
Разрушение
крестцового отдела |
Психогенная
эрекция у 25% больных обеспечивается пояснично—грудными сегментами |
Возможны,
если наступает эрекция |
Возможен |
|
Перерыв
спинного мозга в области верхних грудных или шейных |
Наблюдается
почти всегда (рефлекторная) |
Практически
никогда не встречается |
Невозможен |
Рефлексы, приводящие к эрекции,
замыкаются на уровне II—IV крестцовых сегментов спинного мозга. При полном
поперечном повреждении спинного мозга выше крестцового отдела эрекции у мужчин сохраняется.
Она может отчасти сохраняться и при полном разрушении крестцовых сегментов
спинного мозга, что определяется функцией преганглионарных симпатических
волокон, происходящих из последних грудных и первых поясничных сегментов
спинного мозга (рис. 14.4). Тела постганглионарных симпатических нейронов
располагаются в нижнем брыжеечном ганглий или в узелках вблизи полового члена
(см. разд. 5.1.7).
Во время сексуального возбуждения
нервные импульсы воздействуют в конечном счете на парасимпатические центры
внутренностного тазового нерва и приводят к тому, что тонически сокращенные в
обычных условиях артерии полового члена расширяются и волна крови вливается в
сосудистую сеть пещеристых тел, заполняя и растягивая их. По окончании полового
акта или в ситуациях, неблагоприятных для половых проявлений, возбуждаются
симпатические центры спинного мозга и через нижнее брыжеечное симпатическое
сплетение воздействуют на сосудистую сеть полового члена, подавляя эрекцию.
Эмиссия и эякуляция. Эмиссия и эякуляция —
это рефлекторные процессы, являющиеся заключительной, кульминационной стадией
коитуса. До начала эмиссии секрет яичек в результате перистальтических
сокращений их придатков накапливается в семявыносящих протоках. С наступлением
эмиссии под влиянием сокращений семявыносящих протоков, обладающих сильной
гладкой мускулатурой, секрет яичек проходит в предстательную часть
мочеиспускательного канала, где он смешивается с секретами предстательной
железы, семенных пузырьков, желез уретры. Образовавшаяся здесь смесь секретов
носит название спермы. Сперматозоиды (от греч. sperma — семя, zoon — живое существо и eidos — вид) открыты А. Левенгуком (1677)
и свое название семенные животные получили за подвижность.
Эмиссионный рефлекс (см. рис. 14.4)
возникает в результате раздражения механорецепторов головки полового члена при
трении его. о стенку влагалища во время фрикций. Сенсорные соматические и
парасимпатические волокна в составе полового и тазового нервов достигают
крестцовых сегментов спинного мозга, симпатические — грудопоясничных. С
возрастанием возбуждения во время коитуса происходит активация эфферентных
симпатических путей рефлекса, что сопровождается сокращением внутреннего
сфинктера мочевого пузыря. Закрытие сфинктера препятствует попаданию спермы в
мочевой пузырь.
Вслед за эмиссией начинается
выбрасывание семени — эякуляция. Толчком к эякуляции служит скопление
спермы, которое раздражает чувствительные окончания симпатической и
парасимпатической природы заднего отдела уретры. В этом рефлексе участвуют
также сенсорные пути от придатков яичек, семявыносящих протоков, семенных
пузырьков. Эфферентный путь рефлекса проходит через крестцовые отделы спинного
мозга. Под влиянием эфферентных импульсов происходят тонические и клонические
сокращения луковично—губчатых и седалищно—пещеристых мышц, которые окружают
проксимальные участки пещеристых и губчатых тел. При этом сокращаются также
мышцы тазового дна. В результате массивных ритмических сокращений происходит
интенсивное толчкообразное выбрасывание спермы. Момент эякуляции сопровождается
ритмическими сокращениями туловищных соматических мышц и толчкообразными
движениями таза. В целом все это способствует забрасыванию спермы в
проксимальные отделы влагалища и шейку матки. В заключительной части эякуляции возникает оргазм. По времени он
совпадает с сильными сокращениями соматической мускулатуры.
Время наступления эякуляции при половом акте зависит от многих факторов
— интенсивности половой жизни, темперамента и прочих условий. Количество
спермы, выделяющееся при эякуляции, в среднем составляет 2 мл. Оно также
зависит от частоты повторных половых сношений, эмоционального состояния и
других факторов.
Объем
эякулята у хозяйственно полезных животных различен: 50—100 мл у жеребца, 4—5 мл
у быка, 200—400 мл у хряка, 1—2 мл у барана.
14.8.2. Половые рефлексы у женщин
Половое возбуждение у женщин
сопровождается рядом изменений наружных и внутренних половых органов и прежде
всего увеличением размеров наружных половых органов в результате их усиленного кровоснабжения.
В основе этих изменений лежат два механизма.
Первый связан с раздражением рецепторов половых губ,
преддверия влагалища, а также промежности и лобка. Среди них особое значение
принадлежит клитору, в котором имеется огромное число механорецепторных
приборов (рис. 14.5). Эти
рецепторы возбуждаются в результате прямого раздражения при надавливании,
например корнем полового члена на клитор и другие наружные половые органы.
Возбуждение в этом случае по волокнам полового нерва достигает крестцовых сегментов
спинного мозга. В рефлекторных реакциях участвуют также и висцеральные
афферентные волокна, идущие в составе симпатических стволов. Возбуждение
значительно усиливается при набухании половых органов.
Второй механизм — сугубо психогенного происхождения. Он
связан с функцией головного мозга.
Усиленный приток крови к половым губам, клитору, преддверию
влагалища возникает в результате возбуждения вазоактивных симпатических
волокон, а также сосудорасширяющих парасимпатических эфферентных нейронов
крестцовых сегментов спинного мозга. Эрекция клитора, как и полового члена,
связана с наполнением кровью его пещеристой ткани.
|
|
|
Рис. 14.5 Иннервация половых органов женщины 1 — большая
половая губа, 2 — оргастическая
манжетка, 3 — клитор, 4 —
малая половая губа, 5 —
лобковый симфиз, в — мочевой
пузырь, 7 — маточная труба, 8 —
яичник, 9 — матка, 10 — семяпринимающая полость, 11 — влагалище, 12 —
симпатические нейроны; 13 —
грудные и поясничные сегменты спинного мозга, 14 — крестцовые сегменты спинного мозга, I5 — парасимпатические нейроны, 16 — двигательные нервные волокна к мышцам тазового дна, 17 — прямая кишка; широкой стрелкой
обозначены нисходящие влияния; спинномозговые интернейроны и функциональные
модули метасимпатической нервной системы не показаны. |
Спустя примерно
полминуты после сенсорного или психогенного полового возбуждения через плоский
эпителий влагалища начинается выхождение в его полость слизистой жидкости. Эта
жидкость предназначена для облегчения введения полового члена во влагалище, а
также способствует адекватному раздражению рецепторов полового члена во время
его движения при половом акте. Выхождение жидкости возникает в результате
переполнения стенки влагалища венозной кровью и опосредовано рефлекторными
процессами с участием крестцовых парасимпатических и грудопоясничных
симпатических механизмов.
По мере нарастания полового
возбуждения изменяются и размеры матки. Она увеличивается примерно на 50%.
Меняется и ее положение в полости таза: матка постепенно поднимается и
отклоняется от задней стенки влагалища. В результате такого движения во
внутренней части влагалища вблизи шейки матки возникает полость для приема
семени — семяпринимающая полость. Увеличение размеров матки, ее движения
и приподнятие связаны с усиленным кровенаполнением не только самой матки, но и
области таза. В приподнятии матки определенное значение имеет сокращение
гладкомышечных элементов ее связочного аппарата.
При сильном рефлекторном или
психогенном половом возбуждении большие половые губы истончаются и, наполняясь
венозной кровью, расходятся в стороны, обнажая малые половые губы. В это же
время из—за интенсивного притока крови величина малых половых губ возрастает в
2—3 раза, так что они выпячиваются за пределы больших половых губ, значительно
удлиняя тем самым канал влагалища. По мере возрастания возбуждения аналогичные
изменения возникают и в клиторе. Набухая, он увеличивается. Эта ретракция
клитора сопровождается и его подтягиванием вперед к лобковому симфизу.
С возрастанием возбуждения во время
полового акта в нижней трети влагалища появляется оргастическая манжетка
(оргастическая платформа) (рис. 14.5). Она возникает из—за местного застоя
крови и представляет собой сужение стенки влагалища по всему его периметру. В
момент оргазма и в зависимости от его интенсивности может возникнуть несколько
таких сокращений манжетки.
Оргазм. Оргазм представляет собой ощущение
полового наслаждения в момент завершения полового акта, охватывающее в целом
весь организм. Биологическое значение оргазма состоит прежде всего в том, что
он является побудительным фактором к совершению нормального полового акта с
целью зачатия новой жизни и продления вида.
Оргазм возникает в результате
суммации возбуждений эрогенных зон (головка полового члена, клитор, вульва,
влагалище), наступающих во время фрикций. Ощущение, достигая кульминации,
завершается у мужчин эякуляцией, а у женщин — выталкиванием слизистой пробки,
плотно закрывающей маточный зев, сокращением оргастической манжетки и матки. В
зависимости от интенсивности оргазма может происходить до 15 сокращений
оргастической манжетки. Они представляют собой аналог эмиссии и эякуляции у
мужчин. Регулярные сокращения матки начинаются от ее дна и охватывают все тело.
В основе возникновения сокращений оргастической манжетки и матки лежат
симпатические рефлекторные механизмы.
Наряду с реакциями половых органов,
возникающими под влиянием автономной нервной системы, во время оргазма
возбуждение охватывает висцеральные органы и особенно интенсивно центральную
нервную систему. В этот момент частота сердечных сокращений может достигать 180
ударов/мин, диастолическое давление повыситься на 20—40 мм рт. ст.,
систолическое — на 30—100 мм рт. ст., частота дыхания — до 40 вдохов в минуту,
возникают сокращения наружного сфинктера заднего прохода.
В момент оргазма произвольный
контроль за соматической мускулатурой в значительной мере исчезает, возникают
непроизвольные, почти судорожные сокращения брюшных, межреберных и лицевых
мышц. Общие реакции висцеральных органов и особенно интенсивное возбуждение ЦНС
приводят в совокупности к усилению половых ощущений. Особенно оно проявляется у
женщин, часто сопровождаясь подавлением других видов чувствительности.
У мужчин и женщин время наступления и
формы проявления оргазма разнообразны и индивидуальны. Это зависит от
физического и нервно—психического состояния, интенсивности полового влечения и
темперамента, образа жизни, возраста, ситуации и других факторов. У мужчин
половой акт обычно завершается оргазмом.
В отличие от мужчин женщины во время
полового акта могут многократно испытывать оргазм. Если оргазм не наступает, то
стадия спада длится у женщин дольше. Отсутствие оргазма у женщин не
препятствует зачатию. После оргазма наступает спад половой возбудимости,
восстановление которой происходит в разные сроки в зависимости от
индивидуальных особенностей личности.
С наступлением половой зрелости в
половых органах женского организма человека и животных возникают периодические
изменения получившие название циклов. Их регуляция осуществляется
эндокринной системой (см. разд. 6.3.3). Во время каждого цикла
происходит созревание одного или нескольких фолликулов и овуляция — выход из
фолликула зрелой, способной к оплодотворению яйцеклетки. Параллельно с
созреванием фолликула во время цикла возникают изменения в слизистой оболочке
половых органов. Достигая определенного максимального уровня, эти изменения
вновь подвергаются обратному развитию. У разных видов животных частота,
длительность цикла, морфологические проявления сильно различаются, хотя лежащие
в их основе физиологические механизмы являются принципиально одинаковыми.
По частоте возникновения циклов млекопитающие разделяются на
моноэстральных (от новолат. oestrus, estrus — течка) и полиэстральных. К
первым относится большинство диких животных, у которых обычно в течение года
возникают 1—2 цикла. Вторую группу составляют многие хозяйственно—полезные
животные, большинство грызунов и человек. У них в течение года циклы
повторяются многократно.
Тип половой периодичности во многом зависит от биологических
условий размножения. Он может меняться под влиянием ряда факторов внешней
среды, таких как климат, обилие корма и т. д. Например, дикие моноэстральные
животные в неволе могут становиться полиэстральными. При всем разнообразии
половой цикл самок большинства млекопитающих состоит в общем из нескольких фаз
или периодов: предовуляционный, овуляционный, послеовуляционный, покоя.
В предовуляционном, или проэстральном,
периоде происходит увеличение фолликулов, одновременно разрастается эпителий
матки. Во влагалищном секрете появляются ядерные эпителиальные клетки. У многих
моноэстральных животных этот период является периодом подготовки полового
аппарата к половому акту. Предовуляционные изменения возникают под действием
гонадотропного гормона гипофиза на внутрисекреторную функцию яичников, в
результате чего усиливается выработка эстрогенов (см. разд. 6.2.3).
В овуляционном, или астральном,
периоде происходит разрыв фолликула и выход из него зрелой, способной к
оплодотворению яйцеклетки — ооцита. Биологическая надежность
воспроизведения вида у приматов и человека обеспечивается огромным количеством
ооцитов, достигающим в препубертатном возрасте 300 тыс. Однако в каждом
астральном периоде из 10—15 одновременно растущих фолликулов овулирует лишь
один.
Во время овуляции возникает усиление притока крови к
маточным трубам, напряжение их гладкомышечных волокон, сокращение мерцательного
эпителия. Это сопровождается расширением воронки и сумки яйцевода, схватыванием
ими яичника, что обычно способствует у человека попаданию яйцеклетки и
фолликулярной жидкости в яйцевод. Последующие попеременные сокращения мышечных
волокон яйцевода продвигают яйцеклетку в сторону матки. Время прохождения
яйцеклетки по трубе до матки составляет у женщин около 3 сут. По мере
приближения момента овуляции и особенно в период овуляции происходит
перестройка функций половых органов и организма в целом. Эти изменения
возникают под влиянием эстрогенов, образующихся в фолликулах. При значительном
количестве эстрогенов у животных возникает течка и половая охота.
Течка (эструс) представляет собой комплекс сложных структурных и
функциональных изменений, происходящих в репродуктивных органах самки. Во время
течки возникает попеременное сокращение и расслабление мышц матки, ослабляется
мышечное напряжение шейки матки, что делает ее канал проходимым. Под действием
эстрогенов расширяются кровеносные сосуды слизистых оболочек половых путей, они
приобретают малиновую окраску и становятся отечными. В мазке, взятом со стенок
влагалища, обнаруживается огромное количество эпителиальных клеток с хорошо
сохранившимися ядрами. У грызунов происходит сильное ороговение поверхностного
эпителия, который затем слущивается. Поэтому течку у них можно установить по
мазкам слизистой оболочки влагалища. Усиленная функция секреторных клеток
приводит к образованию больших количеств слизи, вытекающей наружу. У приматов и
человека кровотечение во время овуляции отсутствует, менструальные же
кровотечения связаны не с овуляцией, а с отторжением части эндотелия
приблизительно через две недели после овуляции (см. ниже).
Во время течки у самки наступает половая охота. Она
занимает только часть того времени, в течение которого длится течка, и
характеризуется готовностью самки к половому акту. Самка в охоте становится
беспокойной и стремится приблизиться к самцу. При встрече с ним проявляет
готовность к совокуплению, принимает соответствующую позу и др.
Выделившаяся из фолликулов яйцеклетка может быть
оплодотворена. Оплодотворение возникает лишь в том случае, если
спаривание произойдет непосредственно перед овуляцией или сразу же после нее.
Поэтому самки млекопитающих, кроме приматов и человека, склонны спариваться
лишь в определенные периоды продолжительностью в несколько дней. Эти периоды
совпадают с циклическим повышением уровня половых гормонов в крови (см. разд.
6.3.3). Сам процесс спаривания длится у собаки до 45 мин, у свиньи — 5—8, у
лошади — 1,5—2 мин, у жвачных — несколько секунд. У млекопитающих овулировавшие
яйцеклетки сохраняют в маточной трубе способность к оплодотворению относительно
непродолжительное время: у кролика — 6—8 ч, мыши — 15, морской свинки — 20 ч.
Если при спаривании почему—либо не произошло оплодотворения,
то возникает следующая стадия — послеовуляционный период. Он наступает,
когда на месте лопнувшего фолликула вслед за овуляцией образуется желтое тело,
которое развивается из стенок опорожненного фолликула. Фолликулярные клетки
делятся и превращаются в лютеоциты (лютеиновые клетки).
Желтое тело является временной железой, продуцирующей гормон прогестерон.
Через несколько суток оно начинает рассасываться и полость бывшего фолликула
заполняется соединительной тканью. При этом уменьшается, а затем прекращается
выработка прогестерона. В яичниках начинают развиваться новые фолликулы, и
снова возрастает секреция эстрогенных гормонов. Неоплодотворенная яйцеклетка
остается в матке в течение нескольких дней и затем погибает. С исчезновением
желтого тела слизистая оболочка матки подвергается обратному развитию. У человека
и высших обезьян ее целостность нарушается, в результате чего наступает
кровотечение — менструация (от лат. menstrus — ежемесячный).
Появление менструального кровотечения определяется рядом факторов.
После прекращения функции желтого тела резко уменьшается содержание эстрогенов
и прогестерона, что отражается на изменении кровообращения в сосудах слизистой
оболочки матки.
Застой крови в сосудах и замедление кровотока приводит к
повышению давления внутри сосудов, стенки их разрываются и начинается
кровотечение. Одновременно возникают тонические сокращения маточной
мускулатуры, приводящие к отторжению слизистой оболочки. Отторгнувшиеся части
вместе с кровью выводятся наружу.
Вслед за послеовуляционным периодом наступает период межовуляционного
покоя (диэструс). В это время фолликулы относительно малы, слизистая
оболочка матки бледная, эпителий мало развит, секрет влагалища содержит
лейкоциты. Период покоя переходит в предовуляционный период следующего цикла.
Длительность полового цикла у одного и того же животного
варьирует значительно меньше, чем у разных животных одного и того же вида.
Например, у коровы она составляет 19—21 сут, кобылы — 20—22, буйволицы — 21—30,
овцы — 10—14, свиньи — 20—21, козы — 17 сут, собаки — 3—6 мес. Каждый период
полового цикла характеризуется изменениями не только в половых органах самок,
но и в организме в целом.
У женщин половой цикл принято называть менструальным
циклом. Условно его определяют от первого дня наступившей менструации до
первого дня следующей менструации. Длительность менструального цикла у женщин
детородного возраста (18—45 лет) колеблется в пределах от 21 до 35 сут.
Нормальным считается менструальный цикл, продолжающийся 28 сут, так как при
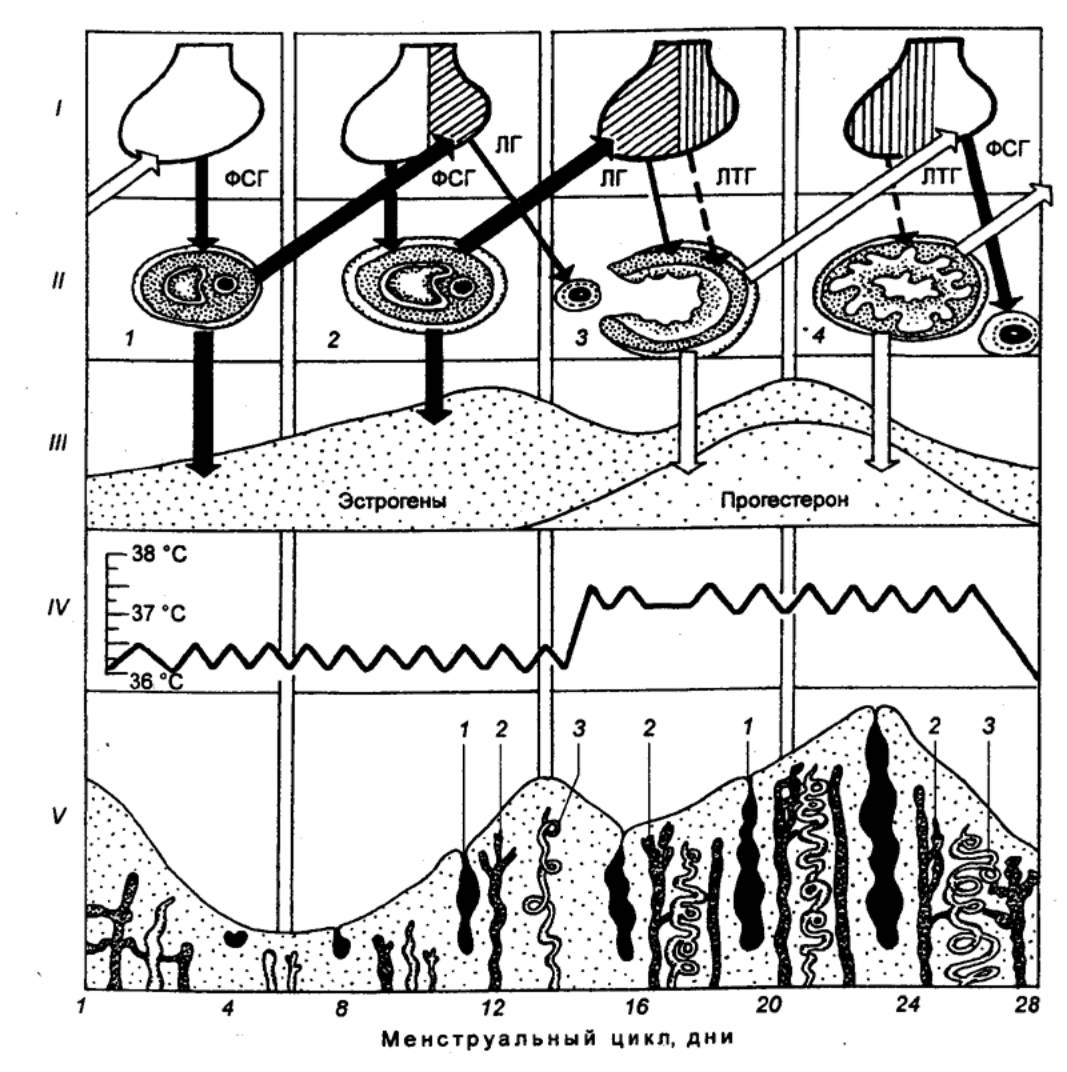
этом наблюдается особенно строгая периодичность циклических изменений (рис.
14.6). Начинается менструальный цикл в периоде полового созревания (11—16 лет)
и продолжается до 45—50 лет. Первая менструация появляется у девочек, живущих в
Европейской части России, в среднем в 12 лет и 9 мес. Средняя продолжительность
менструаций 2—3 сут.
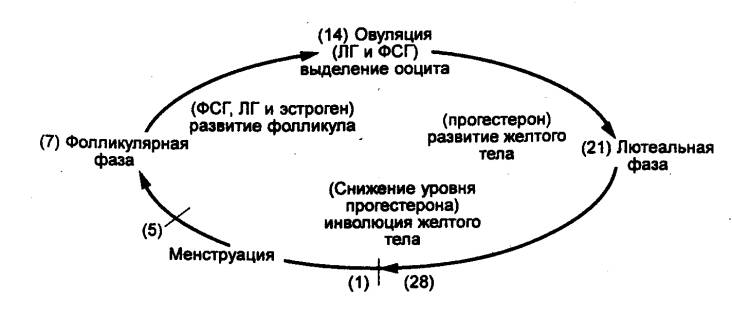
В менструальном цикле принято рассматривать четыре стадии
(рис. 14.7) — менструация, фолликулярная фаза, овуляция, лютеальная фаза. В
событиях, происходящих во время менструального цикла, участвуют яичники
(овариальный, или яичниковый, цикл) и матка (маточный цикл). Оба они
регулируются гормонами яичников, секреция которых в свою очередь находится под
контролем гипофизарных гонадотропинов.
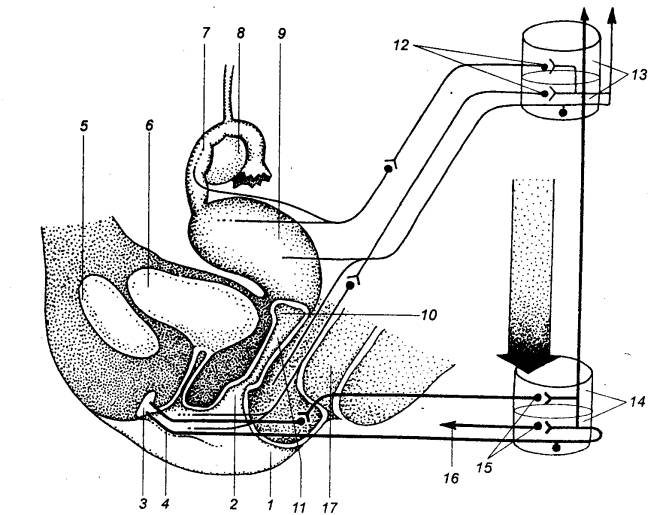
Овариальный цикл у взрослой женщины начинается, как известно, с развития
нескольких первичных фолликулов (рис. 14.8). Они содержат ооциты первого
порядка. Из них лишь один продолжает расти и превращается в зрелый
|
|
|
Рис. 14.6 Схема 28—дневного менструального цикла I — циклические изменения в последовательность секреции
гормонов гипофиза; незаштрихованное изображение гипофиза соответствует
превосходящей по сравнению с другими гормонами секреции ФСГ, косая штриховка
— секреции ЛГ, прямая штриховка — секреции ЛТГ; II — циклические изменения в яичнике: 1,2 — фаза созревания фолликула, 3, 4 — овуляция, фаза желтого тела и начало созревания нового
фолликула; III —
циклические изменения выделения гормонов созревающим фолликулом и желтым
телом; 1У — двухфазная кривая базальной температуры: в фазу
созревания фолликула она не превышает 37 °С, с момента овуляции и в фазу
желтого тела базальная температура выше 37 оС; V — циклические изменения в
эндометрии, соответственно, фазам менструального цикла: 1 — железы (изменение их размера и
секреции), 2 — вены, 3 —
артерии (увеличение их длины и извилистости); стрелками показано
взаимодействие гормонов гипофиза и яичника. |
|
|
|
Рис. 14.7 Главные фазы менструального цикла (цифры в
скобках — дни, на которые приходится данная фаза) |
|
|
|
Рис. 14.8 Схема яичника женщины
(в разрезе) Показаны и обозначены цифрами последовательные стадии
развития граафова пузырька, овуляции, образования желтого тела и его
инволюции. Наблюдать все эти стадии одновременно в действительности нельзя. |
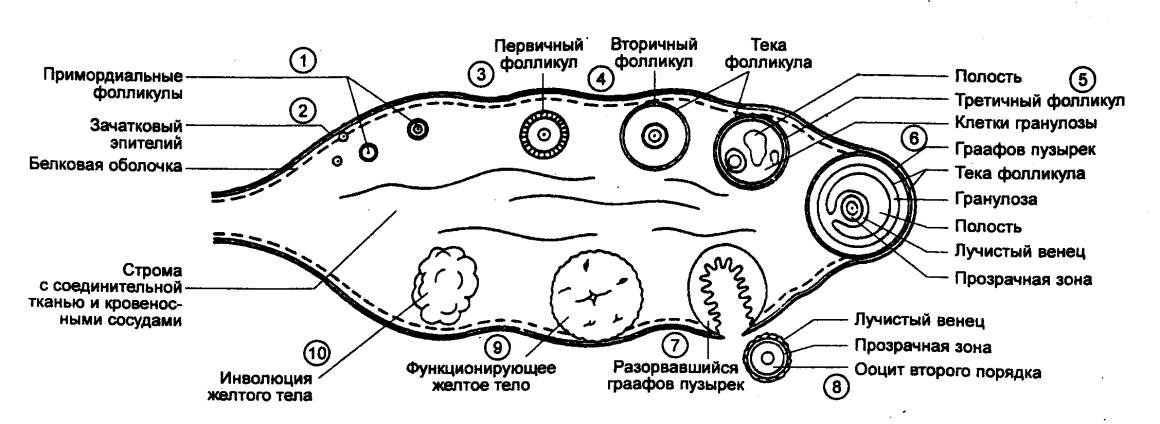
везикулярный яичниковый фолликул,
часто называемый граафовым пузырьком. Граафов пузырек достигает примерно 1 см в
диаметре и выступает над поверхностью яичника в виде бугорка. В момент овуляции
ооцит отделяется от стенки лопнувшего фолликула, выходит в брюшную полость и
попадает в маточную трубу. Как правило, каждый месяц освобождается только один
ооцит одним из яичников, так что овуляция происходит поочередно то в одном, то
в другом яичнике.
|
|
|
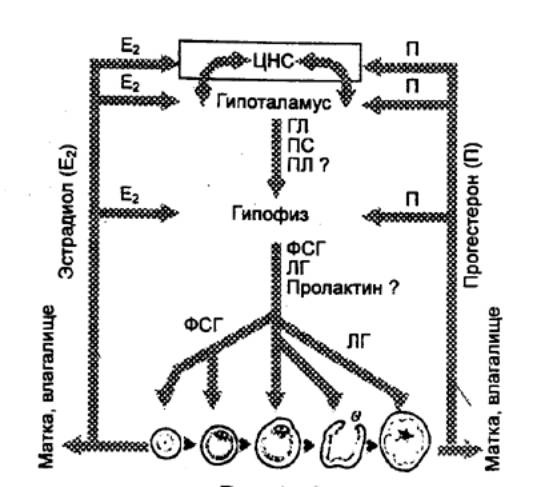
Рис. 14.9 Регуляторная
система гипоталамус—гипофиз—яичники ФСГ — фолликулостимулирующий
гормон, ЛГ — лютеинизирующий гормон, ПЛ — пролактин, ПС — пролактостатин, ГЛ
— гонадолиберин. |
Маточный цикл также делится на фазы — менструальную, пролиферативную и
секреторную. Фазы связаны со структурными и функциональными изменениями
эндометрия. В менструальной фазе происходит отторжение эпителиального
слоя эндометрия и выведение его остатков вместе с кровью. Пролиферативная
фаза совпадает с фолликулярной фазой овариального цикла и состоит в быстрой
пролиферации клеток эндометрия, приводящих к его утолщению. Во время секреторной
фазы выделяемый желтым телом прогестерон стимулирует секрецию слизи
трубчатыми железами. Слизь подготавливает выстилку матки для возможного
принятия оплодотворенного яйца.
Гормональная регуляция овариального и маточного циклов
осуществляется системой гипоталамус—гипофиз—яичники (рис. 14.9). Общая схема
этой регуляции сейчас представляется следующим образом. Декапептид гонад
гонадолиберин, высвобождаемый гипоталамическими нейронами, стимулирует секрецию
гонадотропинов — фоликулостимулирующего гормона (ФСГ) и лютеинизирующего
гормона (ЛГ). ФСГ индуцирует созревание фолликулов. Как уже указывалось, у
женщин одновременно созревает только один из них (крайне редко — два).
Созревающий фолликул образует все возрастающие количества эстрадиола,
вызывающего пролиферацию эндометрия. Достигнув достаточно высокой концентрации,
эстрадиол по механизму положительной обратной связи усиливает в середине
менструального цикла выброс гонадолиберина и повышает чувствительность к нему
гипофизарных клеток, секретирующих ФСГ и ЛГ. В результате возникает пик выброса
этих гормонов в середине цикла. ЛГ вызывает овуляцию и лютеинизацию гранулезных
клеток фолликула. Последние под его влиянием начинают секретировать
прогестерон. Стероиды, эстрадиол и прогестерон действуют на гипоталамус и
гипофиз по механизму отрицательной обратной связи, подавляя во второй половине
цикла секрецию ФСГ и ЛГ. Оба гормона действуют также на высшие отделы ЦНС,
поэтому у животных половое влечение (либидо) в значительной мере зависит от
стадии цикла.
В семенниках и добавочных половых железах самцов не
наблюдается периодических изменений, подобных тем, что происходят в половых
органах самок. Периодические изменения в семенниках, сезонная активация
сперматогенеза имеются только у животных со строго сезонным размножением
(например, лисица, соболь и др.). Так, начиная с конца марта у самцов лисиц происходит
резкая дегенерация семенников и через некоторое время они по своему строению
напоминают половые железы неполовозрелых щенков. Восстановление половых желез
происходит у них в декабре, а в конце января—февраля у лисиц начинается
спаривание.
Оплодотворением называют слияние сперматозоида с
яйцеклеткой, приводящее к образованию зиготы, способной расти, развиваться и
давать начало новому организму. Во время оплодотворения ядерный материал
мужской и женской половых клеток объединяется, что приводит к объединению
отцовских и материнских генов, восстановлению диплоидного набора хромосом.
Для многих беспозвоночных животных и большинства рыб,
обитающих и размножающихся в воде, присуще наружное осеменение яйца.
Сперматозоиды у них выбрасываются в воду, где и наступает оплодотворение. У
животных, размножающихся на суше, осеменение внутреннее. При помощи
совокупительного органа самец вводит сперму непосредственно в половые пути
самки.
Первыми позвоночными, разрешившими проблему оплодотворения и
развития на суше, были рептилии. Совершенно очевидно, что на суше нельзя просто
выбрасывать гаметы в окружающую среду. Стало быть, первым условием для полного
перехода к наземной жизни было введение мужских гамет в тело самки, т. е.
внутреннее оплодотворение. И поскольку при этом вероятность слияния гамет резко
возрастает, то и число их может быть значительно уменьшено.
Различают два типа внутреннего осеменения: влагалищный
и маточный. При первом эякулят попадает во влагалище, оттуда сперматозоиды,
активно двигаясь, переходят в шейку матки. При втором типе сперма вводится
непосредственно в матку. Влагалищный тип осеменения свойствен жвачным,
некоторым грызунам и приматам, маточный — лошади, свинье, собаке и другим
животным.
Тип осеменения зависит от устройства матки, ее шейки, от
степени развития добавочных половых желез у самцов. У животных с маточным типом
осеменения матка очень длинная, а у самцов добавочные железы хорошо развиты и
объем эякулята значителен, что является важным биологическим приспособлением,
так как при большом объеме жидкости возможно проталкивание сперматозоидов на
сравнительно значительные расстояния.
У животных с влагалищным типом осеменения объем эякулята
небольшой. Анатомическое строение и физико—химические условия в шейке матки
делают ее местом, пригодным для длительного хранения сперматозоидов. В шейку
матки они попадают из влагалища.
У человека эякулят вводится во влагалище. Его объем
составляет 2—5 мл и содержит в 1 мл от 30 до 100 млн. сперматозоидов. Однако из
них через канал шейки матки в ее полость проникает всего несколько миллионов, а
верхней части маточной трубы достигает лишь около 100 сперматозоидов.
Оставшиеся во влагалище сперматозоиды не могут существовать там долго из—за
кислой среды (рН 5,7), хотя некоторая защита в этом случае и обеспечивается
щелочными свойствами эякулята. В полости матки условия выживания спермиев также
не столь благоприятны, но уже по другой причине. Здесь играет главную роль
высокая фагоцитарная активность лейкоцитов. Далее, одним из препятствий в
продвижении сперматозоидов к яичнику являются и трудности механического
перемещения в маточно—трубной области. Все это в целом имеет и свою
положительную сторону, предотвращая попадание в маточные трубы ослабленных или
необычных половых клеток. Выжившие сперматозоиды уже через 10—20 мин после
полового акта могут достичь ампулы маточной трубы. Такое быстрое продвижение
одна лишь подвижность сперматозоидов обеспечить не может. Продвижению
способствует целый ряд факторов, к числу которых относятся мышечные сокращения
влагалища, сокращения миометрия, цилиарные движения, перистальтические
сокращения и ток жидкости в маточных трубах. В отдельных случаях сперматозоиды
проходят всю длину маточной трубы и оплодотворяют яйцеклетку непосредственно
после овуляции, до ее попадания в воронку яйцевода. В таких случаях
прикрепление зародыша может произойти к яичнику или стенке брюшной полости, что
приводит к развитию внематочной беременности.
Период, в течение которого
сперматозоиды в половых путях самки сохраняют способность к оплодотворению,
относительно непродолжителен: у мыши — 6 ч, у морской свинки — 22, у кролика —
до 36 ч. У женщин в половых путях сперматозоиды сохраняют способность к
оплодотворению на протяжении 2 сут. У животных имеются исключения. Так, у
некоторых летучих мышей спаривание происходит осенью, а овуляция яйцеклеток и
их оплодотворение осуществляется только весной. Таким образом, сперматозоиды у
них сохраняют способность к оплодотворению на протяжении нескольких месяцев.
Существует два типа оплодотворения:
моно— и полиспермия. При моноспермии благодаря особым механизмам в
яйцеклетку проникает лишь один сперматозоид. Этот вид оплодотворения широко
распространен у животных с наружным осеменением, хотя свойствен также и
млекопитающим. При полиспермии в яйцеклетку проникает до нескольких
десятков сперматозоидов, однако ядро только одного из них соединяется с женским
пронуклеусом, остальные посредством специальных механизмов ооплазмы исключаются
из развития. Этот тип оплодотворения присущ животным с внутренним осеменением
(членистоногие, моллюски, хордовые). Он, по—видимому, является эволюционно
вторичным. Слияние гамет и последующее преобразование у животных с разными
типами оплодотворения протекает в основном одинаково и по единому плану.
Число овулирующих фолликулов зависит
от вида млекопитающего. На этот процесс могут оказывать влияние многие факторы
окружающей среды. В обычных условиях у человека, как правило, овуляции
достигает лишь один фолликул, хотя посредством гонадотропных гормонов можно вызвать
суперовуляцию. Механизмы, с помощью которых из всей массы развивающихся
фолликулов выбирается один определенный, изучены мало. Процесс этот происходит
в первые несколько дней цикла, сразу после начала менструации. Выбранный
фолликул начинает расти и дифференцироваться, опережая остальные и становясь
доминантным. Параллельно с развитием доминантного фолликула остальные
подвергаются атрезии (обратному развитию).
|
|
|
Рис.
14.10 Строение зрелого спермия
человека |
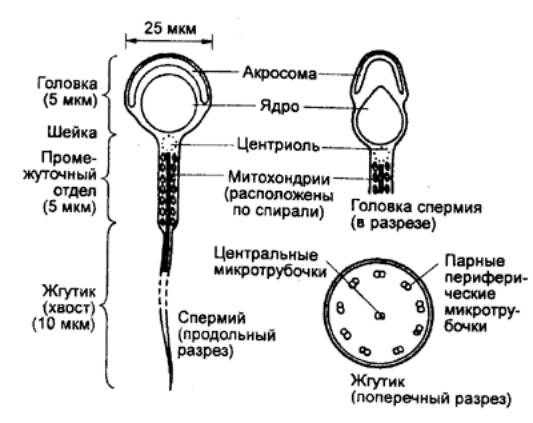
Оболочка яйцеклетки насекомых и
многих рыб содержит отверстия, через которые сперматозоиды проникают в
яйцеклетку. У млекопитающих такие отверстия отсутствуют, и, чтобы достигнуть
ооплазмы, сперматозоиды должны пройти через толщу оболочек. Для этого у сперматозоида
имеются специальные органеллы — акросомы, располагающиеся на вершине его
головки (рис. 14.10). Погрузившись в ооплазму, головка сперматозоида
ориентируется в глубь яйцеклетки и претерпевает сложные изменения. При этом его
ядро постепенно преобразуется в мужской пронуклеус. Эти преобразования идут
одновременно с формированием женского пронуклеуса. Мужской и женский
пронуклеусы сближаются в центре яйцеклетки и затем сливаются, образуя единое ядро
зиготы. На этом завершается процесс оплодотворения.
Оплодотворение
включает следующие процессы: узнавание яйцеклетки сперматозоидом; регуляцию
поступления сперматозоида внутрь яйцеклетки, предотвращение полиспермии;
окончание второго мейотического деления; образование мужского и женского
пронуклеусов, начало клеточного деления.
Для процесса узнавания характерно
несколько механизмов, и прежде всего известно, что в качестве рецепторов для
сперматозоидов выступают гликопротеиды прозрачной оболочки яйцеклетки.
Рецепторы эти высокоспециализированы и видоспецифичны. Это полностью исключает
какие—либо межвидовые слияния половых клеток.
Поступление сперматозоида внутрь
яйцеклетки начинается с возникновения большого числа контактов между
плазматической мембраной и акросомальной мембраной сперматозоида. В результате
взаимодействия появляются пузырьки с протеолитическими ферментами. Эти ферменты
как раз растворяют матрикс фолликулярных клеток и прозрачную оболочку. В
образовавшийся благодаря ферментативному воздействию канал в прозрачной
оболочке сперматозоид проникает, используя пропульсивную силу хвоста.
Предотвращение полиспермии также
достигается посредством ряда механизмов, главный из которых состоит в том, что
тотчас же после пенетрации (проникновения) первого сперматозоида происходит
практически мгновенная деполяризация мембраны яйцеклетки, переходящая в стойкий
блок (процесс детально изучен у морских ежей). Полный блок возникает в
результате активации кортикальных гранул, представляющих собой лизосомные
органеллы, содержащие протеолитические ферменты. Содержимое гранул изливается в
околоклеточное пространство и проникает в прозрачную оболочку. В результате
рецепторы к сперматозоидам инактивируются, сама же прозрачная оболочка
становится плотной и недоступной для последующих вмешательств мужских половых
клеток.
Произошедшее слияние сперматозоида и
яйцеклетки запускает входящий кальциевый ионный ток и освобождение кальция из
внутриклеточных депо, осуществляя тем самым активацию оплодотворенной
яйцеклетки (зиготы). Через ряд промежуточных механизмов зигота вступает в
первое митотическое деление. Для возникновения стадии образования двух клеток
бывает необходимо от 24 до 36 ч.
Образовавшаяся после оплодотворения
зигота постепенно продвигается по направлению к матке и выходит в нее через
несколько суток. В течение 2—3 дней она находится в полости матки во взвешенном
состоянии. Питание осуществляется за счет находящейся там жидкости.
Прикрепление (имплантация) же зиготы к стенке матки происходит лишь на 6—7—й
день после овуляции. В этот период эндометрий стенки матки в результате
воздействия эстрогенов и прогестерона оказывается подготовленным к процессу
имплантации.
На овуляцию, оплодотворение и
имплантацию может направленно влиять ряд средств и способов контрацепции
(предохранение от зачатия. Об этом также необходимо здесь кратко заметить, т.
к. последнему процессу принадлежит немалое практическое значение.
Методы контрацепции принято делить на две группы —
обратимые и необратимые. К обратимым относятся методы, основанные на прерывании
контакта между сперматозоидом и яйцеклеткой. Это: 1 — прерывание полового акта
(извлечение полового члена до наступления эякуляции), 2 — ритмический, 3 —
барьерный. Низкая эффективность первого связана с весьма вероятной потерей
контроля мужчиной, а попадания небольшой порции сперматозоидов еще до эякуляции
оказывается вполне достаточным для наступления беременности. При ритмическом
методе главным условием является воздержание в так называемый «опасный период»,
который в течение 3—4 дней
предшествует овуляции и столько же дней
продолжается после нее. Ненадежность этого метода обьясняется тем, что овуляция
является индивидуальным процессом каждой женщины и даже при самом тщательном
контроле не всегда удается точно определить время ее наступления. Барьерные
методы предусматривают применение специальных предметов — цервикальных
колпачков, диафрагм, спиралей и презервативов. Первые закрывают вход в канал
шейки матки и подбираются гинекологом. Самым распространенным и надежным
средством предупреждения беременности являются мужские презервативы, применяемые
в сочетании со спермицидными средствами. Считается также, что презервативы
защищают от заболеваний, передающихся половым путем — венерических болезней и
СПИДа.
К необратимым методам предупреждения
беременности относятся вазэктомия (операция иссечения всего семявыводящего
протока), предотвращающая у мужчин попадание сперматозоидов в эякулят, и
перевязка маточных труб у женщин. Вполне понятно, что оба названных метода
готовятся на основании предварительных операций и могут быть использованы лишь в
крайних случаях. Ясно также, что после таких операций восстановить функцию
семяпроводов и маточных труб практически невозможно.
Есть еще и гормональные способы
контрацепции. Действие их основано на предотвращении овуляции в результате
подавления секреции гонадотропных гормонов в середине цикла, а также и влиянии
их на половые органы. Действующим началом в этом случае являются зстрогены и
прогестерон.
Беременностью называется физиологический процесс,
при котором из оплодотворенной яйцеклетки в женском организме развивается плод.
В эмбриональном развитии организма млекопитающих выделяют
три периода: зародышевый, во время которого образуются бластоциста,
трофэктодерма (трофобласт), происходит закладка зародышевых листков и плодных
оболочек;
предплодный, являющийся переходным от
зародышевого к плодному и характеризующийся закладкой всех органов, и плодный,
начинающийся от конца первой трети беременности и завершающийся родами. В этом
периоде продолжается рост и развитие всех органов, а также формирование плода в
особь, сходную с родительским организмом.
Во время зародышевого периода оплодотворения яйцеклетка,
продвигаясь по маточной трубе в сторону тела матки, делится митотическим путем
на дочерние малодифференцированные клетки — бластомеры. Таким образом,
одноклеточный зародыш — зигота — превращается в многоклеточный зародыш — морулу,
состоящую из 8—12 клеток. В моруле различают два слоя клеток: светлые и темные.
Светлые клетки, образующие наружный слой, играют роль в питании зародыша и
носят название трофэктодермы. Из темных, центральных клеток (внутренней
клеточной массы) — эмбриобласта — развивается сам зародыш.
С попаданием зародыша из маточной трубы в полость матки
размножение клеток резко ускоряется и зародыш приобретает вид полого пузырька —
бластоцисты.
У приматов и человека с помощью протеолитических ферментов
зародыш разрушает прилежащий участок слизистой оболочки матки, целиком
погружается в ее толщу, где и продолжает развиваться. Бластоциста увеличивается
в размере, ее оболочка разрывается, и образуется желточный пузырь. Он
содержит белковую жидкость, которая возникает из секрета слизистой оболочки
матки. Параллельно с этим начинается образование плодных оболочек —
амниона (водной), аллантоиса (мочевой) и хориона (сосудистой).
Амнион образует околоплодный пузырь. Функцией его эпителия является секреция
околоплодных вод, определяющих важнейшие условия развития плода, а также
выведение продуктов его обмена в околоплодную жидкость. Вторая оболочка — аллантоис
— представляет собой пузырь, облегающий амнион. Третья оболочка — хорион —
вместе с аллантоисом составляет аллантохорион. У млекопитающих хорион тесно
связан со слизистой оболочкой матки, образуя детское место, или плаценту (рис.
14.11).
Плацента является временным органом,
имеющимся только у плацентарных млекопитающих. Строго говоря, это единственный
орган, состоящий из клеток, имеющих происхождение из двух разных организмов —
матери и плода.
Во время беременности плацента
обеспечивает связь плода с организмом матери. У разных млекопитающих в
зависимости от формы и расположения, а также отношения ворсинок хориона к
слизистой оболочке матки различают четыре типа плаценты: эпителиохориальную,
десмохориальную, эндотелиохориальную и гемохориальную.
Эпителиохориальный тип плаценты устроен наиболее просто.
Хорион в ней только прилегает к эпителию слизистой оболочки матки, и его
ворсинки располагаются в криптах слизистой оболочки. В этом случае источником
питания плода является эмбриотроф — секрет, выделяемый железами матки. Этот
тип плаценты характерен для непарнокопытных животных.
При десмохориальном типе
плацента состоит из отдельных долек, называемых котиледонами, в которых
сгруппированы сильно ветвящиеся ворсинки. Соответственно котиледонам на слизистой
оболочке матки развиваются материнские плаценты — карункулы. Они имеют
грибовидную форму с множеством углублений на поверхности. В котиледонах
ворсинки хориона внедряются в слизистую оболочку матки, разрушая ее эпителий,
врастают в соединительнотканный слой. Благодаря этому связь между оболочкой
плода и слизистой оболочкой матки становится более прочной. Такой тип плаценты
присущ жвачным животным.
Эндотелиохориальный тип плаценты имеет форму муфты или
пояса. Ворсинки хориона разрушают эпителий слизистой оболочки матки, строму и
мышечные стенки ее сосудов. Эпителий хориона в этом случае непосредственно
прилегает к эндотелию сосудов матки. Этот тип плаценты встречается у плотоядных
и грызунов.
Гемохориальный тип плаценты является наиболее
совершенным типом. Изменение слизистой оболочки матки происходит более глубоко
— хорион разрушает все ее элементы, в том числе и эндотелий сосудов, так что
сложно ветвящиеся ворсинки хориона оказываются не только погруженными в кровь
матери, но и свободно в ней плавают. Этот тип плаценты характерен для обезьян и
человека (см. рис. 14.11).
Посредством плаценты осуществляется обмен
веществами между матерью и плодом, при этом выполняются функции газообмена,
трофическая, эндокринная, выделительная, защитная. Плацента обладает
антигенными и иммуногенными свойствами. Она не только объединяет, но и
разделяет генетически неоднородные организмы матери и плода, предотвращая
возникновение иммунологического конфликта.
Обмен газами между кровью матери и плода происходит
путем их диффузии через ворсинки хориона. У человека площадь ворсинок,
приходящихся на
|
|
|
Рис.
14.11 Взаимоотношения между
пупочными кровеносными сосудами, капиллярами ворсинок хориона и наполненными
кровью лакунами в плаценте человека |
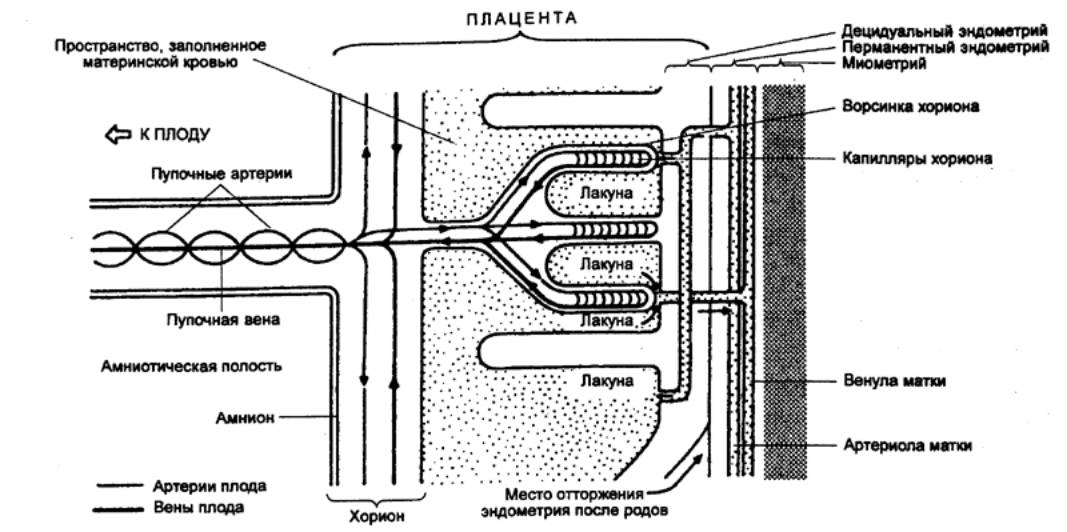
1 кг массы плода, более чем в 3,5
раза превышает площадь поверхности легочных альвеол, соответствующей 1 кг массы
взрослого человека. Движение 02
и СО3 происходит здесь по градиенту концентрации этих газов.
Питание плода через хорион происходит строго
избирательно. Оно осуществляется в результате сложных процессов ферментативного
расщепления белков, углеводов, жиров и синтеза питательных веществ в
соответствии с потребностями развивающегося плода. В плаценте содержатся ферменты,
расщепляющие и синтезирующие белки, имеется значительное количество ферментов и
стероидных гормонов, влияющих на углеводный обмен. В цитоплазме клеток плаценты
есть липиды и липолитические ферменты, принимающие участие в синтезе гормонов.
Уровень минеральных веществ — кальция, железа, фосфора, меди, цинка, марганца,
кобальта и других микроэлементов — в крови плода превышает их уровень в
материнском организме. Считают, что многие из этих веществ не только
используются для непосредственных потребностей плода, но и расходуются
новорожденным. Плацента содержит также большое количество разнообразных
витаминов. Накапливая их, она осуществляет и регуляцию поступления этих веществ
к плоду. В плаценте имеются факторы свертывания крови (см. разд. 7.7). Они способствуют
прекращению кровотечений после родов.
Плацента обладает эндокринной
функцией. В ней происходят процессы синтеза, секреции и превращения
гормонов (см. разд. 6.3.3). Эти гормоны определяют развитие приспособительных
изменений в организме матери, необходимых для роста и развития плода,
осуществления родового акта и лактационной функции.
В плаценте образуются
«гипоталамический» рилизинг—фактор, прогестерон и эстрогены (гормоны яичников).
Их функция направлена на сохранение и прогрессирование беременности, а также
изменение эндокринных органов матери. Плацента синтезирует целый ряд гормонов,
к числу которых относятся: характерный для беременности гормон белковой
природы, плацентарный лактоген человека, хорионический гонадотропин,
пролактин, гестагены. Как паракринные агенты функционируют образующиеся в
плаценте некоторые пептиды, включающие кортикотропин—рилизинг—фактор и
инсулиноподобный фактор роста.
В плаценте имеются
видоспецифические, групповые, тканево—органные антигены гистосовместимости,
ответственные за индукцию реакции трансплантационного иммунитета. Ткани амниона
и хориона, несмотря на единое зародышевое происхождение, качественно различны в
антигенном отношении. Этот факт не имеет пока объяснения, однако в таком
различии заложен определенный биологический смысл. Иммунологическая
«инертность» плодовой части плаценты к материнскому организму является важным
фактором взаимозащиты матери и плода. Она предохраняет от развития
иммуноконфликтных реакций.
Плацента обладает способностью
регулировать проникновение различных веществ из крови матери к плоду и в
обратном направлении. Эта совокупность морфологических и функциональных
особенностей получила название плацентарного барьера. В функциональном
отношении он сходен с гематоэнцефалическим барьером, но отличается тем, что
участвует в обмене веществ двух организмов, обладающих значительной
самостоятельностью. Морфологическим субстратом плацентарного барьера служат
эпителиальный покров ворсинок и эндотелий располагающихся в них капилляров.
Факторами, обусловливающими проницаемость, являются состояние и функциональная
активность мембран.
Проницаемость плацентарного барьера
в определенной мере зависит от молекулярной массы веществ. Вещества с
молекулярной массой до 600 Да проходят его сравнительно легко. Для веществ,
имеющих молекулярную массу выше 1000 Да, плацента почти не проницаема. Вместе с
тем некоторые биологически активные вещества типа вазопрессина, тироксина с
высокой молекулярной массой относительно быстро проникают через плацентарный
барьер.
Барьерная функция плаценты при
переходе от матери к плоду белков, углеводов, жиров, витаминов, электролитов,
кислорода и других необходимых веществ регулируется механизмами, возникающими в
процессе фило— и онтогенеза. По отношению к веществам, случайно попавшим в
организм матери или введенным в ее кровь, барьерные функции плаценты выражены в
меньшей степени или вовсе отсутствуют. Через плаценту проникают алкоголь,
наркотики, никотин, ртуть, мышьяк, гемолитические яды, многие токсические
вещества. Сейчас считают доказанным переход к плоду возбудителей инфекционных
болезней — вирусов и микробов. Плацентарный барьер ограничивает, но не
препятствует проникновению к плоду многих лекарственных веществ, таких как
антибиотики, сульфаниламиды, барбитураты, анальгетики, гликозиды, гормоны.
У человека с момента имплантации и
до 9—й недели внутриутробного развития развивающийся организм носит название зародыша,
или эмбриона, начиная с 9—й недели и до рождения — плода. Беременность
у женщин в большинстве случаев продолжается около 9 месяцев, и роды обычно
происходят через 280 дней, или 40 недель, считая с первого дня последней
менструации. Продолжительность беременности у разных видов животных различна
(табл. 14.2).
В конце I месяца беременности у
женщин зародыш имплантирован в слизистую оболочку матки, в нем происходит
возникновение зачатков органов и плодовых оболочек. В конце II месяца
дифференцирована головка и туловище, III месяца — конечности. В конце IV месяца
различим пол, лицо почти сформировано, начинается окостенение черепа. В связи с
образованием мышечной системы нарастает активность движений конечностей,
появляются слабые дыхательные движения. В конце V месяца движения настолько
активны что ощущаются матерью, может быть прослушано сердцебиение, в кишечнике
формируется первородный кал (меконий). В конце VI месяца созревают
Таблица 14.2
Продолжительность беременности у различных видов
хозяйственно полезных животных, сут
|
Вид животных |
В среднем |
Колебания |
|
Лошади |
340 |
307—412 |
|
Ослы Коровы |
380 285 |
360—390 240—310 |
|
Верблюды |
365 |
335—395 |
|
Северные олени |
225 |
195—243 |
|
Свиньи |
114 |
110—140 |
|
Овцы, козы |
150 |
140—160 |
|
Собаки |
62 |
59—65 |
|
Лисицы |
51 |
51—54 |
|
Кошки |
58 |
55—60 |
|
Норки |
42 |
37—80 |
|
Соболи |
275 |
250—300 |
|
Кролики |
30 |
28—33 |
внутренние органы. В конце VII
месяца масса плода достигает 1 кг, длина — 35 см. Из—за плохого развития подкожной
клетчатки плод имеет «старческий» вид, все тело покрыто пушковыми волосками. В
конце VIII месяца плод жизнеспособен, но нуждается в условиях внутриутробного
развития. В конце IX месяца плод приобретает признаки зрелости, «старческий»
вид утрачивается, кожа становится розовой. Доношенный плод (т. е. имеющий
внутриутробный возраст 40 недель) имеет длину не менее 47 см и массу не менее
2500 г.
Становление функций органов и систем плода начинается еще в
эмбриональном периоде. На самых ранних стадиях развития происходит образование ферментных
систем, катализирующих синтез веществ, которые необходимы для жизни и
развития плода. На II месяце в печени развивается система гемопоэза. Сердце начинает
функционировать к концу 4—й недели, и после этого возникает движение крови в
сосудах эмбриона, возрастающее с увеличением массы плода. Волокна
блуждающего и симпатического нервов появляются на 7—й неделе, после
чего частота сердечных сокращений у плода становится в два раза чаще, чем на
ранних стадиях развития. В конце беременности частота сердечных сокращений
плода составляет 120—140 ударов/мин. Кровообращение плода имеет свои
особенности (см. разд. 8.9). Артериальное давление постепенно повышается от
39/21 мм рт. ст. на V месяце внутриутробной жизни до 80/45 мм рт. ст. при
рождении.
У зрелого плода структура дыхательной системы такая
же, как и у взрослого человека. Одновременно с развитием дыхательной системы
развиваются и регулирующие ее образования в ЦНС. Плод непрерывно совершает
нерегулярные дыхательные движения. Они осуществляются при закрытой голосовой
щели, поэтому массового захвата околоплодных вод не происходит, а небольшие их
количества, попадающие в дыхательные пути, всасываются.
Нервная система закладывается в конце I месяца беременности, в конце II месяца
обнаруживаются первичные ядра и проводящие пути. У зрелого плода регистрируются
биотоки головного мозга, что свидетельствует о включении коры больших полушарий
головного мозга в регуляцию рефлекторной деятельности. Органы чувств у зрелого
плода развиваются не одинаково. Слух и болевая чувствительность недостаточны,
зрение ограничивается светоощущением осязание и термочувствительность развиты
хорошо. Все эндокринные железы начинают функционировать к концу 20—й
недели.
Деятельность эндокринной системы плода осуществляется
преимущественно автономно, что и понятно, так как во время внутриутробного
развития в полной мере не задействован ряд органов и систем. Например, почки и
легкие не работают соответственно как органы экскреции и газообмена (их функцию
выполняет плацента), температурные условия не требуют затрат энергии для своего
поддержания, опорно—двигательная система не участвует в осуществлении локомоций
и позных реакций. В целом гормоны в организме плода выполняют те же самые
функции, что и в организме взрослого. Помимо того, они регулируют и некоторые
важные процессы, происходящие исключительно в период внутриутробного развития.
К их числу относятся такие процессы, как дифференцировка пола и начало родовой
деятельности.
14.11.3. Состояние организма матери при беременности
Неотъемлемую часть механизма
развития беременности составляют рефлекторные реакции, возникающие в
результате сложных взаимоотношений между организмом матери и плода. Они
начинаются с возбуждения рецепторов матки. К моменту родов падает возбудимость
коры головного мозга и, наоборот, повышается возбудимость спинного мозга. Это
приводит к усилению спинномозговых рефлексов, повышению рефлекторной и мышечной
возбудимости матки.
Форма, величина, положение и
реактивность матки во время беременности значительно изменяются. Она постепенно
увеличивается в течение всей беременности в результате гипертрофии мышечных
волокон. С увеличением срока беременности в мышце матки и плаценте
накапливается большое количество биогенных аминов, что приводит к резкому
повышению спонтанной возбудимости и энергетической готовности сократительных
белков к выполнению механической функции. Под влиянием вырабатываемых в
плаценте гормонов происходит рост и гиперплазия молочных желез и подготовка их
к лактации (см. разд. 6.2, 14.14). Эта секреция активируется лютеотропным
гормоном гипофиза.
Значительные изменения происходят в
эндокринных железах. С началом беременности овуляция в яичниках прекращается,
желтое тело некоторое время продолжает функционировать. Продуцируемые им
гормоны способствуют нормальному развитию беременности. Мощной эндокринной
железой, в которой интенсивно протекают процессы образования, превращения и
выделения гормонов, является плацента. Диапазон биологического действия
ее гормонов очень широк. Наряду с плацентой в синтезе ряда гормонов участвует
плод. Его метаболическая активность и активность плаценты дополняют
материнскую. Во время беременности вследствие размножения и гипертрофии клеток,
вырабатывающих гормоны, увеличивается передняя доля гипофиза, повышается
образование гонадотропных, тиреотропного, гормонов, кортикотропина.
Супраоптическим и паравентрикулярным ядрами гипоталамуса усиливается
образование окситоцина и вазопрессина. Эти ядра принимают также участие в
секреции аденогипофизом фолликулостимулирующего, лютеотропного, и тиреотропного
гормонов, кортикотропина.
С перестройкой функций эндокринной и
нервной системы при беременности значительным изменениям подвергается обмен
веществ. Возрастают основной обмен и потребление кислорода, заметно
изменяется белковый обмен. В крови беременных повышено содержание жирных
кислот, холестерина и других липидов, которые расходуются на образование тканей
плода. Многие неорганические соединения также задерживаются в организме матери.
Увеличивается усвоение фосфора, необходимого для развития нервной системы и
скелета плода, солей кальция, идущих для построения костей плода, происходит
значительное накопление железа, макроэлементов — К, Na, Mg, Cl, микроэлементов — Со, Сu и др. Эти вещества также переходят
к плоду. Они определяют нормальное течение процессов обмена и роста. В связи с
необходимостью снабжения плода и поддержания интенсивного обмена веществ
возрастает потребность организма матери в витаминах.
Увеличение объема циркулирующей
крови, повышение периферического сопротивления, развитие мощной системы маточно—плацентарного
кровообращения предъявляют повышенные требования к работе сердца и сосудистой
системы беременной. При беременности происходит физиологическая гипертрофия
левого желудочка, возрастание минутного объема, учащение пульса, повышение
сосудистого тонуса. Артериальное давление изменяется незначительно. Объем
циркулирующей крови к концу беременности возрастает на 35—40%, усиливается
кроветворение, увеличивается количество гемоглобина.
Дыхание в последние месяцы
беременности становится несколько чаще и глубже, происходит усиление
газообмена. Усиливается деятельность почек, выделяется больше мочи,
мочеиспускание становится более частым. Под влиянием вырабатываемого в плаценте
релаксина происходят изменения в костно—мышечной системе. Они
характеризуются разрыхлением суставных связок, хряща и суставных оболочек
сочленений тазовых костей. В связи с этим увеличиваются подвижность таза и его
емкость во время родов. Своеобразным изменениям при беременности подвергается
кожа. Довольно часто в коже лица, сосков, белой линии живота наблюдается
коричневая пигментация. Ее появление связано с усиленной продукцией в
надпочечниках близкого к меланину пигмента.
14.11.4. Многоплодная беременность
Многоплодная беременность может
возникнуть либо в результате оплодотворения двух (или более) яйцеклеток, и
тогда развиваются разнояйцовые близнецы, либо при развитии эмбрионов из
одной оплодотворенной яйцеклетки, тогда развиваются однояйцовые близнецы.
Многоплодная беременность у женщин
встречается в 0,4—1,6% случаях наиболее часто при этом бывают двойни.
Каждая оплодотворенная яйцеклетка после внедрения в
слизистую оболочку матки образует свои плодные оболочки, из которых у каждого
плода развивается своя плацента с самостоятельной сетью сосудов. У однояйцовых
близнецов амниотическая оболочка может быть либо отдельной у каждого эмбриона,
и тогда они остаются обособленными, либо общей, тогда соответственно и полость
для обоих зародышей будет общей.
При многоплодной беременности развитие плодов, как правило,
протекает более или менее равномерно, разница в их массе невелика. Расположение
плодов в матке может быть различным, но чаще оба плода находятся в продольном
положении и головном предлежании.
14.11.5. Латентная стадия беременности
У некоторых животных (косули, норки, горностаи, соболи,
полевые мыши, крысы и др.) оплодотворенная яйцеклетка длительное время не
прикрепляется к матке. У соболя, например, сезон спаривания бывает в июле, а
щенение самок происходит в апреле—мае. У этих животных в феврале—марте в матке
обнаруживаются свободные, еще не прикрепленные бластоцисты. Истинная
беременность от начала образования плаценты до родов продолжается в этих
случаях 27—30 дней. Период от оплодотворения до прикрепления зародыша получил
название латентной стадии беременности. Она связывается с появлением в
этот период гормональной неактивности желтых тел. Латентная стадия беременности
представляет собой особую форму биологического приспособления, позволяющую
животному давать потомство в наиболее благоприятный для его кормления и
воспитания сезон года.
14.11.6. Беременность у животных
Беременность является
особенностью отдельных видов низших позвоночных животных, таких как некоторые
рыбы (скаты, акулы, бельдюга и др.), амфибии (альпийская саламандра).
Беременность имеет место и у рептилий (живородящая ящерица, гадюки),
относящихся к высшим позвоночным. Весьма характерно, что у рептилий один и тот
же вид может быть и живородящим, и яйцекладущим. В этом случае все определяют
условия окружающей среды. У живородящих рыб, амфибий и рептилий во время
беременности образуется своеобразная желточная плацента, отдаленно напоминающая
плаценту млекопитающих.
Из высших
млекопитающих непродолжительной и не такой относительно сложной является
беременность у сумчатых. Например, у гигантского кенгуру она длится всего 39
дней. Детеныш рождается недоразвитым и все последующее немалое время (примерно
8 мес) развивается в сумке на животе матери. У высших млекопитающих начальные
стадии развития оплодотворенной яйцеклетки проходят в свободном состоянии,
после чего у китообразных, копытных, хоботных происходит прикрепление зародыша
к стенке матки, а у части насекомоядных, грызунов, человекообразных обезьян —
внедрение в стенку матки. Значительно колеблется и продолжительность
беременности у разных видов — от 19—20 дней у мышей до 660 дней у слонов.
В зависимости от
экологических условий окружения у разных видов животных эмбрионы развиваются не
только по—разному, но и рождаются с разной степенью зрелости (козленок, способный
вскоре после рождения следовать за матерью, и лисенок, остающийся
продолжительное время слепым и беспомощным). Борьбой за существование служит и
такое явление, как плодовитость. У крупных животных (китообразные, мозоленогие)
беременность продолжительная и одноплодная, другие виды, например, копытные и
хищные животные, характеризуются укороченной беременностью с несколькими
плодами в помете. Для грызунов характерна не только многоплодность, но и
многопометность. Это означает, что животное приносит несколько многоплодных
пометов за один сезон размножения.
Роды — физиологический процесс изгнания из
матки через родовые пути плода по достижении им жизнеспособности, а также
плаценты с околоплодными оболочками и околоплодными водами. Родовой акт —
сложный многозвеньевой процесс, возникающий и завершающийся в результате
взаимодействия многих систем организма. В возникновении родов наибольшая роль
принадлежит нейрогуморальной и гормональной системе матери и плодоплацентарному
комплексу (рис. 14.12).
В последней стадии беременности
перед родами в материнском организме наблюдается преобладание тормозных
процессов в высших отделах мозга и
|
|
|
Рис. 14.12 Основные гормональные и нервные механизмы.
связанные средами Цифры 1—7 указывают на возможную
последовательность событий, вызывающих сокращение стенки матки; 8 — афференты, 9 — эфференты |
повышение возбудимости спинного
мозга, что приводит к снижению порога рефлекторной и мышечной возбудимости
матки. При этом усиливаются интероцептивные влияния с половой сферы. Становясь
надпороговыми, эти интероцептивные влияния могут способствовать началу и
развитию родового акта.
Для наступления родов большое значение имеют взаимоотношения
прогестерона и эстрогенов. Пока в организме беременной существует и
поддерживается между ними физиологическое равновесие, сократительной
деятельности матки не происходит. Вырабатываемый желтым телом, а затем
плацентой прогестерон снижает возбудимость маточной мускулатуры, что позволяет
ей растягиваться под давлением плодного пузыря до значительных размеров.
Под влиянием одной из фракций эстрогенов — эстрадиола
— происходит повышение чувствительности матки к медиаторам и особенно к
ацетилхолину, выделяющимся окончаниями моторных нейронов местной сети
метасимпатической нервной системы. Повышение возбудимости матки ведет к
прекращению ее свободного растяжения, повышению внутриматочного давления,
напряжению стенки матки. При этом возрастает чувствительность мышечной оболочки
матки (миометрия) к окситоцину, вызывающему ее сокращения. Окситоцин в большом
количестве накапливается в организме перед родами.
В повышении возбудимости сократительных мышечных
элементов матки наряду с ацетилхолином и окситоцином большое значение
принадлежит серотонину, гистамину, кининовой системе, простагландинам (см.
разд. 5.2). Серотонин, например, обеспечивает проницаемость клеточных мембран
для ионов Са2+, которые активируют ферментативные системы,
необходимые для сокращения мышечной клетки. Эстрогены, серотонин и катехоламины
изменяют соотношение адренергических компонентов, усиливающих функцию α—адренорецепторов и облегчающих высвобождение
прогестерона от связи с белком клеток матки.
Комплекс нейрогуморальных и
эндокринных изменений, происходящих в материнском организме перед родами,
составляет родовую доминанту. В этих условиях большая роль в наступлении
родовой деятельности принадлежит импульсам с интероцепторов матки. Они исходят
от созревшего плода, который начинает усиленно двигаться. По подчревным и
внутренностным тазовым нервам афферентная импульсация достигает спинальных,
супраспинальных и корковых структур мозга (см. рис. 14.5). Когда возбудимость
матки и сила интероцептивного
раздражения достигают определенного предела, мускулатура матки начинает
ритмически сокращаться, появляются регулярные родовые схватки. В течение родов
выделяют три периода: раскрытие шейки матки изгнание плода, последовый
период.
В первом периоде во время схваток, длящихся несколько
часов, происходит сокращение мышечных волокон матки, смещение их относительно
друг друга и растяжение. Волна сокращений распространяется сверху вниз, от
одного рога к другому, затем переходит на тело и далее захватывает нижний
сегмент с убывающей продолжительностью и интенсивностью. С каждой схваткой
возникает укорочение и расширение шейки матки. Раскрытию шейки способствует
плодный пузырь. Во время схваток давление в нем повышается и он смещается в
сторону шейки матки, где встречает меньше препятствий. В результате шейка матки
сглаживается, ее канал превращается в растянутую трубку, через которую возможно
прохождение плода. После раскрытия шейки матки под влиянием повышенного
внутриматочного давления разрывается оболочка плодного пузыря, плодные воды
вытекают, увлажняя поверхность половых путей.
Во втором периоде происходит изгнание плода из матки.
Оно осуществляется за счет потуг — одновременного ритмичного сокращения
мышц матки и брюшного пресса. В результате плод продвигается по родовому каналу
в соответствии с направлением его оси, совершая при этом сгибательные, разгибательные
и вращательные движения.
После рождения пересекают пуповину,
отделяя тем самым новорожденного от последа (рис. 14.13). Пуповина представляет
собой шнуровидное образование, соединяющее плод с плацентой, через сосуды
которого осуществляется плодоплацентарное кровообращение. Пуповина входит в
состав последа. Хищники после родов перегрызают пуповину, у других животных она
обрывается под тяжестью новорожденного.
Третий период родов, завершающий, — последовый. В нем происходит
отслоение плаценты и выделение последа. Послед — совокупность
внезародышевых частей плодного яйца, обеспечивающая двустороннюю связь между
плодом и организмом матери. Послед состоит из плаценты, плодных оболочек и
пуповины. Они связаны в анатомическом и функциональном отношениях и выполняют
сложные функции в течение беременности и родов. После рождения плода
функциональная активность последа угасает и он выделяется из полости матки,
отсюда и его название.
|
|
|
Рис. 14.13 Схема изменений в кровеносной системе,
происходящих при рождении ПП— правое предсердие; ЛП — левое
предсердие; ПЖ — правый желудочек; ЛЖ — левый желудочек. |
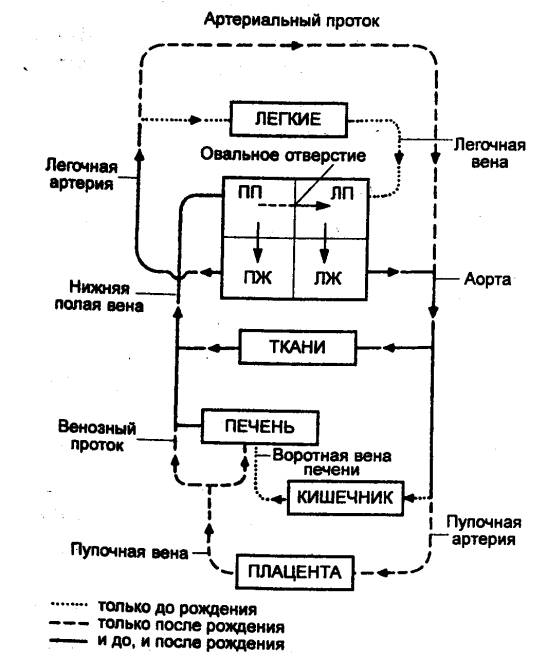
Механизм отслойки плаценты основан на том, что под влиянием сокращения
матки размер плацентарной площадки уменьшается, так как плацента не обладает
способностью к сокращению и начинает отслаиваться. Плацента рождается своей
плодовой поверхностью, которая тянет за собой плодовый мешок со скопившимися в
нем сгустками крови.
У женщин последовый период
начинается с момента рождения плода и завершается выделением последа. У
животных отделение последа происходит через несколько часов после родов, но у
хищных послед выходит вместе с плодом.
Продолжительность родов у первородящих женщин составляет 15—20
ч, у повторнородящих — 10—12 ч. У животных роды чаще всего происходят ночью.
Считают, что в этот период снижается возбудимость коры больших полушарий,
понижается ее тормозное влияние на подкорковые структуры, что способствует
благоприятному течению родовой деятельности. Процесс родов могут затормозить и
даже прервать действия внешних раздражителей, таких как внезапный
шум, яркое освещение и т. д.
По многолетним устоявшимся представлениям сроки наступления
родов находятся под сугубо гормональным контролем, им же и регулируются. Между
тем результаты недавних исследований показали, что у целого ряда млекопитающих
значительную роль в родовом процессе играет плод (см. рис. 14.2). Явление это,
судя по всему, имеет большой адаптивный смысл, так как обеспечивает рождение
плода именно на такой стадии развития, когда плод уже способен к относительно
самостоятельному существованию. Имеется точка зрения, что начальные этапы
изгнания плода вызываются некими еще не установленными окончательно факторами.
Эти факторы якобы заставляют гипоталамус плода стимулировать секрецию
кортикотропина гипофизом. Как свидетельствует одно из современных
представлений, сигналом для начала родов служит «фетальный стресс». Стресс этот
возникает в результате иммунологического отторжения зрелого плода материнским
организмом.
14.13. ОСОБЕННОСТИ РАЗМНОЖЕНИЯ ПТИЦ
Половой процесс птиц имеет ряд
характерных черт. В связи с этим особое значение приобретает физиология
репродуктивного аппарата домашних птиц, таких как куры—несушки, основная
хозяйственно полезная ценность которых связана именно с деятельностью органов
размножения.
Половое созревание яйценосных пород кур завершается к 4—5
мес, более крупных мясных — к 6—8 мес, петушки созревают несколько раньше.
Длительность половой деятельности домашней птицы следующая: петух — до 4 лет,
курица — до 6, индюк — до 3, индейка — до 5, селезень и утка — до 4, гусак — до
5, гусыня — до 8 лет.
14.13.1. Физиология органов размножения самцов
Семенники у самцов расположены в брюшной полости
в области трех последних ребер. В период половой деятельности семенники
становятся крупнее. В их извитых канальцах образуются сперматозоиды. Затем
через семявыносящие канальцы, которые впадают в придаток семенника,
сперматозоиды переходят в семяпровод, открывающийся в стенке клоаки сосочком.
Придатки семенников развиты слабо, придаточных желез у птиц не имеется.
Совокупительного органа у петуха также нет, он вводит семя, прижимая свою
клоаку к клоаке самки. У селезней и гусаков копулятивный орган (половой член)
имеется. Он состоит из фиброзной ткани, образующей пещеристое тело. На его
поверхности спирально проходит семенной желоб, который при эрекции превращается
в канал..
Количество эякулята у
петуха достигает 0,8 мл, количество сперматозоидов в 1 мл составляет 5—7 млрд.,
однако при частом совокуплении число сперматозоидов снижается. По форме
сперматозоиды отличаются от спермиев млекопитающих.
С наступлением линьки половая
активность петуха постепенно ослабевает и наступает состояние полового покоя. В
отличие от яйцеклеток сперматозоиды могут длительное время сохраняться в
яйцеводе кур, не теряя своей оплодотворяющей способности. Например, куры несут
оплодотворенные яйца еще в течение 20 дней после отсадки самцов, у индеек и гусынь этот срок более продолжителен.
У регулярно спаривающихся кур все отделы яйцевода заполнены сперматозоидами.
После спаривания спермин не сразу появляются в воронке яйцевода, где происходит
оплодотворение. Первые оплодотворенные яйца курица несет лишь на третий день.
14.13.2. Физиология органов размножения самок
У птиц функционирует только левый
яичник; правый, хотя и закладывается, вскоре редуцируется. Соответственно у
птиц имеется только один яйцевод, состоящий из воронки, белковой части,
перешейка, «матки» («известковой» части) и влагалища, которое открывается в
клоаку (рис. 14.14). Общая длина яйцевода у кур—несушек достигает 60 см.
Яичник подвешен на брыжейке под
передней долей почки. Его глубокий слой образует сосудистую зону, поверхностный
слой — фолликулы разной величины и зрелости. Они прикреплены на богатых
кровеносными сосудами выростах серозной оболочки. В разгар яйцекладки яичник
напоминает кисть винограда; у курицы—несушки в нем содержится от 500 до 3500
яйцеклеток. Развиваясь и накапливая желток, яйцеклетки превращаются в яичные
желтки, достигающие примерно 35—40 мм в диаметре. На образование желтка таких
размеров требуется около двух недель. На поверхности желтка находится
зародышевый диск, представляющий собой белое пятнышко величиной 1—2 мм. Это и
есть собственно яйцеклетка (рис. 14.15).
В яичнике периодически возникает
овуляция. Она происходит быстро, в течение 1—2 мин. После разрыва фолликулярной
оболочки созревший желток проскальзывает в воронку яйцевода, который при этом
перистальтически сокращается и продвигает его в направлении клоаки. Движению
желтка способствует мерцательный эпителий стенки яйцевода. Прохождение по
верхней части яйцевода до белковой длится около 15—18 мин. В белковой части,
длина которой 30—40 см, формируемое яйцо находится 3—3,5 ч. При продвижении
желток вращается вокруг продольной оси, что происходит благодаря спиральному
расположению слизистой оболочки яйцевода. В процессе вращения он постепенно
окружается четырьмя слоями белка, выделяемого железами яйцевода.
Сначала желток окружается тонким
слоем плотного белка, который вследствие вращения желтка образует на его
полюсах шнурообразные тяжи — градинки (см. рис. 14.15). Благодаря
градинкам желток постоянно удерживается в центре яйца. Здесь, в белковой части
яйцевода откладывается 40—50% белка остальной — в перешейке и матке, где яйцо
находится до 1,5 и 20 ч соответственно. В перешейке начинается образование подскорлупной
оболочки, состоящей из двух листков, которые у тупого конца яйца отходят
друг от друга и образуют в этом месте воздушную камеру — пугу.
С попаданием яйца в матку начинается
образование его известковой оболочки — скорлупы. Она откладывается на
яйце медленно. Во время образования скорлупы происходит усиление кровоснабжения
матки, в результате чего в ее клетки увеличивается поступление солей кальция,
которые выделяются в полость матки. В скорлупе имеется большое число пор, через
которые осуществляется газообмен и испарение воды, что играет важную роль в
развитии зародыша. Большее количество пор расположено на тупом конце яйца, в связи
с чем при хранении яиц в сухом месте увеличиваются размеры пути.
Акт яйцекладки происходит через влагалище — широкое мускулистое
образование, по длине соответствующее длине яйца. Во время снесения яйца матка
опускается, выворачиваются влагалище и клоака. В результате этого яйцо
выскальзывает наружу через верхний край влагалища, не соприкасаясь практически
ни с влагалищем, ни с клоакой. Этот процесс выведения яйца осуществляется
благодаря сокращению мышц клоаки.
Процесс формирования яйца в яйцеводе продолжается от 23 до
30 ч, при этом оплодотворенное яйцо более 80% времени находится в известковой
части яйцевода. У большинства кур—несушек овуляция наступает через 10—30 мин
после снесения предыдущего яйца. Таким образом, овуляция и оплодотворение
происходят с незначительным интервалом, не превышающим 20 мин. Если за это
время оплодотворения не произошло, то яйцо остается неоплодотворенным, даже
если в яйцеводе присутствуют сперматозоиды.
Оплодотворение
осуществляется в воронке яйцевода. В этом случае сперматозоиды проникают в яйцо
через покрывающие его две нежные пленки. Процесс дробления оплодотворенной
яйцеклетки начинается через 4—5 ч после овуляции. Он возникает, когда яйцо
находится в перешейке. При попадании зиготы в матку количество бластомеров
составляет 4—8, а спустя 1 ч после оплодотворения, т. е. к моменту
откладывания, достигает 256. В таком состоянии яйцо более трех недель может
сохранять способность к дальнейшему развитию, если будут созданы
соответствующие температурные условия и влажность.
Процесс яйцекладки у
птиц регулируется нейрогуморальными механизмами. Подобно млекопитающим,
созревание фолликулов в их яичнике стимулируется гонадотропным гормоном
|
|
|
Рис.
14.14 Органы, размножения курицы 1 —
яичник с фолликулами, 2 —
оболочка фолликула, 3 — белковая часть яйцевода, 4 — брыжейка, 5 — сосуды, 6 — перешеек, 7 — матка, 8 — клоака, 9 — прямая кишка, 10
— выходная часть, 11 —
воронка. |
|
|
|
Рис.
14.15 Яйцо курицы 1 —
белковая оболочка, 2,10 —
белый желток, 3 — зародышевый
диск, 4 — желточная оболочка,
5 — известковая скорлупа, б — внутренний слой скорлуповой оболочки, 7 —
наружный слой скорлуповой оболочки, 8
— воздушная камера, 9 —
градинка, 11 — желтый желток. |
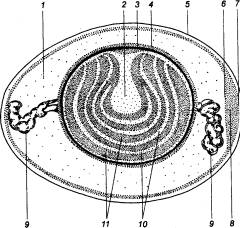
передней доли гипофиза. Еще один гормон гипофиза — лютеинизирующий
— контролирует процесс овуляции. Наряду с этим ряд гормонов образуется и в
самом яичнике. Это эстрин, или фолликулярный гормон, синтезирующийся в
клетках фолликулов, и прогестерон, или гормон желтого тела.
Под влиянием эстрина происходят увеличение яйцевода и
стимуляция деятельности его желез. Повышая метаболизм, эстрин регулирует его
таким образом, чтобы обеспечить синтез и доставку необходимых для образования
яйца веществ. Прогестерон синтезируется в овулировавшем фолликуле яичника. Считают,
что этот гормон регулирует время откладывания яйца.
В естественных условиях яйцекладка у птиц, как
правило, бывает весной. Однако у кур благодаря систематическому отбору на
яйценосность удается получить почти непрерывную яйцекладку в течение всего
года. В этом случае одним из наиболее важных факторов является длина светового
дня. Освещенность стимулирует функцию гипоталамо—гипофизарной системы, что
неизменно отражается на яйцекладке. Отрицательно влияет на яйценоскость линька
с ее наступлением яйцекладка прекращается. Прекращается она и 'при очень
холодной температуре. Внешние раздражители также оказывают влияние на
яйцекладку. Если спугнуть несущуюся курицу, то процесс яйцекладки может
значительно задержаться или даже произойти на следующий день. Испуг несушки
может не только оттянуть яйцекладку, но при этом ухудшить качество яйца. В нем
при этом появляются кровавые точки.
Развитие зародыша у птиц осуществляется при помощи
инкубации. Различают естественную инкубацию — развитие зародыша под
наседкой (насиживание) и искусственную инкубацию — развитие зародыша в
инкубаторе. Наблюдения над насиживанием показывают, что лучшее развитие
зародышей в яйцах происходит при перемежающейся температуре 37—40 оС.
Этот факт и учитывают при создании режима в инкубаторах. При искусственной
инкубации продолжительность эмбрионального развития составляет у кур 20—21 сут, уток и индеек — 27—28, гусей — 29—30
сут.
В первые
5—6 сут инкубации эмбрион развивается наиболее интенсивно В этот период его
питание происходит за счет желтка, который разжижается в результате
ферментативного воздействия, а также из перемещающейся в желток жидкой части
белковой оболочки. С 12 сут у куриных эмбрионов наступает интенсивное
расходование белков, которые полностью используются к моменту вылупления
цыпленка. Неусвоенная часть желтка вместе со стенкой желточного мешка
втягивается перед вылуплением цыпленка в его брюшную полость где и
переваривается.
14.14. ЛАКТАЦИЯ
Вскармливание потомства
молоком матери является одним из важнейших проявлений половой функции, а также
биологических приспособлений у млекопитающих — высших позвоночных животных и
человека. Их анатомическим признаком служат молочные железы, функция которых
состоит в образовании и выведении молока из молочных желез т е лактации.
Лактацией называется помимо того и период времени, на протяжении которого
происходит выделение молока. В целом объем секретируемого молока и
продолжительность его выделения у всех видов млекопитающих рассчитан на
обеспечение пищей новорожденного потомства. Исключение
составляют высокопродуктивные сельскохозяйственные животные (коровы, козы и
др.), удои которых во много раз
превосходят потребности собственного потомства и являются основой для развития
молочной промышленности.
Во время
беременности в молочных железах происходит ряд структурных и функциональных
изменений, подготавливающих железы к отделению молока. начинающемуся
непосредственно после родов. Изменения эти, так же как и процессы,
поддерживающие молочные железы в состоянии готовности к лактации,
контролируются эндокринными факторами на протяжении беременности и
рефлекторными влияниями в период вскармливания потомства.
Строение
молочных желез. Молочные железы
являются кожными железами с экзокринным типом секреции, генетически связанными
с потовыми железами. И хотя у разных животных они имеют разную форму, по—разному
расположены, выделяют молоко неодинакового состава, тем не менее все они имеют
единый план строения.
Железы представляют собой
симметричные образования. У хищных, грызунов и других они расположены
вентрально по сторонам от средней линии тела, у парнокопытных (коровы, овцы) и
непарнокопытных (лошади) — в паховой области, у полуобезьян одна пара на груди и
нередко еще одна пара на животе. У летучих мышей и обезьян одна пара на груди.
У многоплодных форм железы лежат вдоль всей брюшной поверхности тела, число их
может достигать более 20, хотя чаще наблюдается 1—8 пар желез и сосков. Размеры
сосков, их форма зависят от видовой принадлежности. У клоачных млекопитающих
(утконос, ехидна) соски вообще отсутствуют. У ластоногих китообразных соски
скрыты в кожных складках и не выступают наружу даже после родов. .
У женщин молочные, или
грудные, железы располагаются на передней поверхности грудной клетки на
уровне от III до VI или VII ребер. Форма, размеры, положение желез зависят от
возраста женщины, степени развития ее половых органов, от периода беременности,
менструального цикла и индивидуальных особенностей.
Молочные железы у однопроходных
состоят из трубчатых мешков с покрывающими их снаружи гладкомышечными клетками.
Мешки открываются в сумки волос на специальном железистом поле. Более сложное
строение желез у живородящих млекопитающих. Их железы альвеолярного типа, а
железистое поле представлено ограниченной областью в виде выпячивания — соска.
В зависимости от характера выводных протоков принято различать истинные соски и
ложные. На вершине первых открываются многочисленные выводные протоки желез. Этот
тип характерен для большинства сумчатых, обезьян и человека. Ложные
характеризуются тем, что внутри соска имеется общий выводной канал,
открывающийся одним отверстием. Такой тип характерен для копытных и хищных.
|
|
|
Рис. 14.16 Упрощенная схема строения молочной железы 1 — грудная мышца, 2
— альвеолы молочной железы, 3 — млечный проток, 4 — млечные синусы, 5 — отверстия
протоков на соске, 6 — пигментированный
участок (околососковый кружок), 7 — поддерживающая строма, 8 — жировая ткань. |
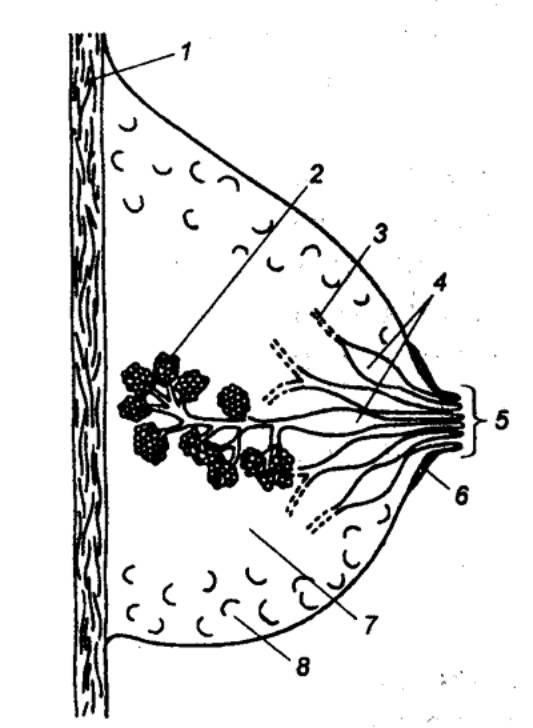
Молочная железа является сложно
организованным образованием. В ней различают собственно железистые структуры,
паренхиму и строму, представленную соединительной тканью. У половозрелой
женщины молочная железа состоит из 15—25 обособленных долей (рис.
14.16). Доли отделены друг от друга прослойками рыхлой соединительной ткани с
жировыми вкраплениями в которых проходят сосудисто—нервные пучки. Каждая доля
представляет собой сложную альвеолярную железку с самостоятельным млечным
протоком. По ходу проток расширяется и образует млечный синус, в
котором накапливается образовавшееся в альвеолах молоко. Проток открывается
самостоятельным отверстием на вершине соска.
В свою очередь каждая доля состоит из более мелких долек
молочной железы, которые образованы многократно ветвящимися млечными
протоками. Протоки эти вне периода лактации заканчиваются слепыми трубочками на
которых в период лактации образуются альвеолы. Стенки альвеол выстланы
однослойным кубическим эпителием, кнаружи от которого располагаются корзинчатые
миоэпителиоциты. Во время лактации эпителиальные клетки трансформируются в
крупные секретирующие клетки — лактоциты, вершины которых обращены в
просвет альвеолы. Характерно, что цитоплазма клеток буквально нафарширована
митохондриями, а у вершины клетки сконцентрированы капельки жира, белковые
гранулы и вакуоли. Это и понятно, так как секреция молока осуществляется по
апокриновому типу посредством отшнуровывания клеточных верхушек вместе с
накопленными включениями. Последующее формирование молока продолжается в
полости альвеолы. Необходимо заметить, что, во—первых, альвеолы выделяют секрет
не одновременно, и, во—вторых, секреторный цикл лактоцитов многократно
повторяется. В период наполнения секретом лактоциты имеют цилиндрическую или
кубическую форму, а после его выведения становятся плоскими. Альвеолы окружены
слоем сократительной миоэпителиальной ткани, участвующей в выведении молока
Увеличение молочных желез во время беременности
связано с развитием их секреторных областей под влиянием эстрогенов,
прогестерона, кортикостероидов, гормона роста, пролактина, плацентарного
лактогена. Однако на всем протяжении беременности наличие в крови прогестерона
подавляет образование молока. При родах, когда уровень прогестерона снижается,
подавление
действия пролактина прекращается и он
стимулирует альвеолы к секреции молока.
Секреция молока. Полный цикл лактации включает развитие железы (маммогенез),
возникновение секреции молока после родов (лактогенез), развитие и поддержание
выделения молока (лактопоэз).
У
млекопитающих особое значение для подготовки и поддержания процесса лактации
имеет пролактин, участвующий также в регуляции широкого спектра процессов, в
том числе роста организма. Гормон имеет много периферических органов—мишеней,
включая молочную железу, печень, надпочечники половые железы, однако ни один
гормон периферического органа не выполняет функции обратной связи для клеток,
синтезирующих пролактин. Уровень базальной секреции пролактина у мужчин и
женщин одинаков. Его секреция существенно повышается во время беременности в
ответ на действие эстрогенов
и поддерживается во
время лактации за счет рефлекторной стимуляции при грудном кормлении.
Молочные железы начинают развиваться у человека на 6—й
неделе внутриутробной жизни. Сосок же формируется в течение первых двух лет
после рождения ребенка. У мужчин на этом развитие молочных желез
приостанавливается. По гистологическому строению они сходны с железами
новорожденных. у женщин же развитие желез продолжается, и к 17—летнему возрасту
оно практически завершается, хотя до наступления беременности в состоянии желез
и возникают изменения, связанные с менструальным циклом. Полным развитием с
образованием альвеол молочная железа характеризуется лишь при беременности и
лактации. В этот период она увеличивается в размерах, расширяются млечные
протоки и вокруг них возникает густая сеть кровеносных сосудов.
После родов в течение первых 2—3
дней лактации выделяется так называемое молозиво — более густой секрет,
нежели зрелое молоко, клейкой консистенции, желтого цвета. Цвет молозива
зависит от. окраски входящего в него жира, богатого каротиноидами. По своему
составу молозиво ближе к тканям организма новорожденного, чем зрелое грудное
молоко. Набор аминокислот молозива занимает промежуточное место между белковыми
фракциями зрелого молока и фракциями крови. По—видимому, это один из
механизмов, способствующий адаптации организма новорожденного в период перехода
от плацентарного питания к питанию грудным молоком. В молозивный период
лактации отмечено повышение концентрации иммуноглобулинов всех классов.
Особенно это касается наиболее биологически активного класса IgA, которые синтезируются в клетках
молочной железы. Молозиво богато и гормонами, особенно кортикостероидами. Это
приобретает особое значение в первые дни жизни новорожденного, так как функции
ряда органов и систем его организма являются еще незрелыми, а активный
иммунитет пока находится в стадии становления. Следовательно, естественное
вскармливание новорожденных с первых моментов появления на свет обеспечивает
местную пассивную иммунологическую защиту его пищеварительного тракта.
Количество зрелого грудного молока у
женщин—рожениц постепенно возрастает и достигает максимальных величин к 8—9—й
неделе. После этого оно, как правило, остается стабильным и составляет 1—1,5 л
в сутки.
В процессе лактогенеза и лактопоэза
эпителиальные клетки молочной железы проходят несколько последовательных стадий
секреторного цикла. Во—первых, начинается поглощение из тканевой жидкости предшественников
молока и последующее накопление этих предшественников в клетке. Во—вторых,
возникает собственный внутриэпителиальный синтез сложных специфических молекул
секрета. В—третьих, образуются гранулы или капли секрета, куда включается
синтезируемый продукт. В—четвертых, осуществляется внутриклеточный транспорт
гранул к апикальной поверхности клетки. В—пятых, происходит выход секрета в
просвет альвеолы. С завершением этого цикла окончательный состав молока еще не
сформирован, наступает следующий этап, называемый «созреванием» молока. Суть
его состоит в реабсорбции некоторых компонентов секрета, сбалансировании
углеводного и минерального состава. Процесс созревания молока осуществляется в
емкостной системе железы.
Расширение и пролиферация млечных ходов,
а также значительное развитие альвеол молочной железы, т. е. стимуляция
лактогенеза происходит под влиянием эстрогенов, прогестерона, пролактина,
соматотропного гормона, лактогенина и других гормонов. Следует иметь в виду,
что полный комплекс лактационных гормонов пока что не выяснен. Однако
считается, что снижение уровня эстрогенов и прогестерона после родов
стимулирует секрецию гипофизарного пролактина и, по—видимому, других гормонов
лактогенного комплекса. В свою очередь выделение пролактина контролируется
гипоталамусом, в котором вырабатывается особый пролактин—рилизинг фактор.
Особая роль в регуляции лактации
отводится окситоцину, стимулирующему отделение молока. Нейроны,
секретирующие окситоцин, сочетают свойства нервной и эндокринной клетки одновременно.
Как классические нейроны, они имеют на своей поверхности множество синапсов.
Мембранный потенциал клетки
|
|
|
Рис.
14.17 Регуляция молоковыведения СО,
ПВ — супраоптическое и паравентрикулярное нейросекреторные ядра гипоталамуса
соответственно. АГ — аденогипофиз, НГ — нейрогипофиз, МЭ — миоэпителий
альвеол, А — альвеола. |
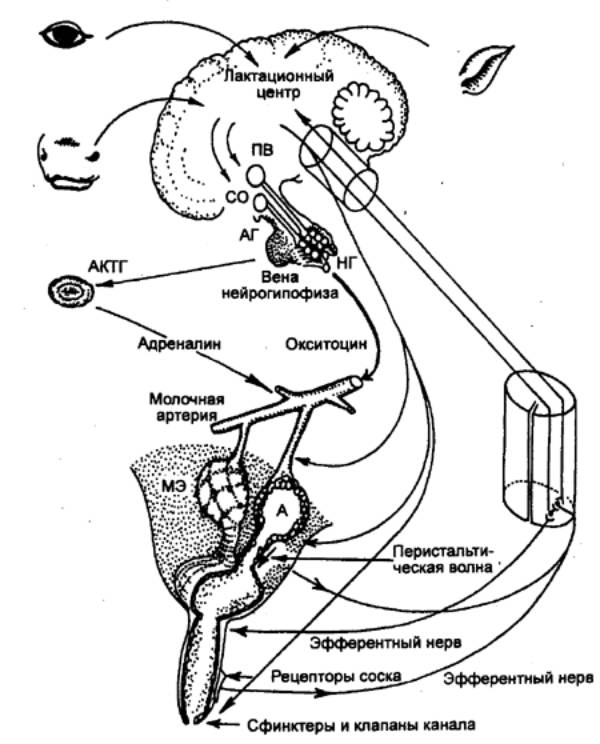
может меняться в широких пределах — от
деполяризации до гиперполяризации. Как только возникший потенциал действия
достигает аксонных терминалей в задней доле гипофиза, он открывает кальциевые
каналы, по которым соответствующие ионы по градиенту концентрации поступают в
клетку. Повышение концентрации внутриклеточного кальция вызывает движение
секреторных гранул, содержащих окситоцин, по направлению к клеточной мембране и
выброс содержимого из клетки. В отличие от типичного нейрона, но аналогично
типичной эндокринной клетке продукт секреции — окситоцин — выделяется не в
синаптическую щель, а во внеклеточное пространство. Отсюда посредством диффузии
окситоцин попадает в капилляры, общий кровоток, к органам—мишеням.
Выведение молока из молочной железы связано с простой
рефлекторной реакцией: когда младенец сосет грудь, он стимулирует рецепторы
соска. Возникающие при этом импульсы через спинной мозг
направляются в гипоталамическую
область. Гипоталамус секретирует окситоцин, выделяющийся через заднюю долю
гипофиза. Окситоцин вызывает сокращение миоэпителиальной ткани, окружающей
альвеолы, и молоко через протоки и синусы выталкивается из сосков (рис. 14.17).
Молоко является сложной биологической
жидкостью, содержащей пищевые вещества в оптимальных соотношениях. Оно служит
наилучшей пищей для ребенка первого года жизни, обеспечивая его полноценное
развитие.
Белки женского молока относятся к
идеальным, их биологическая ценность достигает 100%, 18 белковых фракций
идентичны белкам сыворотки крови. В женском молоке присутствуют четыре класса
иммуноглобулинов — A, G, М, D. Из них наибольшее значение принадлежит глобулину А.
Благодаря низкой протеолитической активности желудочного сока детей первых
месяцев жизни и присутствию ингибитора трипсина в пищеварительном тракте
ребенка не происходит разрушения иммуноглобулинов, что обеспечивает
устойчивость детей этого возраста к инфекционным заболеваниям.
Количество жира в женском и коровьем молоке почти одинаково
(3,5—3,8%), однако жир грудного молока усваивается несравненно лучше (до 95%).
Более высокое содержание ненасыщенных жирных кислот оказывает сберегающее
действие на белок пищи, повышает его усвояемость, способствует проявлению
физиологического действия витаминов, особенно Bi, С и повышает устойчивость
организма к инфекциям. Основным углеводом молока является лактоза. В грудном
молоке ее количество составляет 7,0—7,5%, в коровьем — 4,0—4,5%. Содержащийся в
лактозе моносахарид галактоза в течение первых недель жизни используется
для образования галактоцереброзидов. В грудном молоке содержится также
оптимальное количество минеральных солей, соотношение концентраций которых
является гарантией их наилучшей утилизации.
Молоко синтезируется из метаболитов,
содержащихся в крови: лактоза из глюкозы под действием фермента
лактозосинтетазы, белки — из аминокислот жиры — из жирных кислот, глицерола и
ацетата. Одного только материнского молока достаточно, чтобы младенец ежедневно
прибавлял в весе 25—30 г.
Молочная железа выполняет и функцию
избирательного барьера для ряда лекарственных веществ, принимаемых матерью.
Между тем алкоголь, никотин, кофеин, йод, ацетилсалициловая кислота, некоторые
снотворные ^наркотические вещества и т. д. могут легко переходить в молоко, что
необходимо учитывать в период лактации.
В естественных условиях уровень
секреции молока определяется не только потребностями потомства, но и скоростью
опорожнения железы; особенно отчетливо это выступает у диких видов. У коров при
10—месячной лактации максимальная величина молокообразовательного процесса
приходится на 6—7 неделю лактации, у овец — на 3 неделю. Общее количество
секретируемого молока наиболее высоко у крупных животных. Например, некоторые
киты выделяют его в сутки до 200—250 л. Однако большая интенсивность молочной
продукции характерна для мелких видов млекопитающих. Если исходить из расчета
на единицу массы тела, крыса, например, производит молока в 13 раз больше
коровы.
С окончанием лактации в молочной железе происходит ряд
структурных перестроек и она подвергается функциональной инволюции. В
климактерическом периоде у женщин в связи со снижением функции яичников и, как
следствие, падением уровня эстрогенных гормонов и повышением секреции
фолликулостимулирующего гормона гипофиза постепенно наступает инволюция
секреторных структур железы. Происходит исчезновение альвеол, спадение млечных
протоков и сморщивание железистых долек, замещение железистой ткани фиброзной и
жировой тканями, железа уменьшается в размерах.
В условиях лактации, как правило, происходит
подавление овуляции. Таким образом, женщина находится в состоянии
физиологической контрацепции до тех пор, пока не восстановятся гормональные
изменения, вызванные беременностью и лактацией.
Физиологическое значение репродуктивной системы
сводится к двум основным функциям. Первая обеспечивает образование половых
клеток — овоцитов и сперматозоидов, слияние которых при оплодотворении дает
начало развитию зародыша. Эту функцию часто обозначают как генеративную.
Вторая функция — эндокринная. Она состоит в продукции половых гормонов,
которые оказывают влияние на все процессы, связанные с размножением, начиная с
дифференцировки половых признаков и кончая возникновением беременности,
развитием плода, выращиванием потомства.
У обоих полов репродуктивная система состоит из половых
желез или гонад, и органов полового пути. В мужском организме к последним
относятся семявыносящие пути, предстательная железа, семенные пузырьки, половой
член; в женском — яйцеводы, матка и влагалище, а также тесно связанные с
половой системой молочные железы.
Размножение возможно только с наступлением полового созревания
организма. Созревание состоит в том, что половые органы достигают
полного развития, а в половых железах начинают вырабатываться половые гормоны и
половые клетки. Вся совокупность процессов, обеспечивающих размножение, носит
циклический характер. На определенном этапе развития у созревающего организма
появляются доминанты полового сближения, оплодотворения, беременности, родовая
доминанта. Весь период завершается фазой лактации, обеспечивающей дальнейшее
развитие нового организма, подготавливая его к следующему биологическому циклу.
Мужские половые клетки — сперматозоиды. — образуются
в семенных канальцах. Они открыты в 1677 г. А. Левенгуком. Сперматозоиды
образуются непрерывно и выводятся наружу при половом акте. Сформировавшись в
семенных канальцах, они затем переходят в придаток семенника, где пребывают
длительное время. Здесь сперматозоиды подготавливаются к продвижению на
значительные расстояния, к длительному пребыванию в половых путях самки. Смесь
секретов семенников и их придатков, а также семенных пузырьков, предстательной
железы, бульбоуретральных желез и желез мочеиспускательного канала носит
название спермы.
Яйцеклетки млекопитающих образуются в фолликулах яичников. Впервые их
обнаружил и описал русский ученый академик К. М. Бэр в 1827 г. Каждая
яйцеклетка окружена слоем фолликулярного эпителия, продуцирующего эстрогенные
гормоны. Постепенно фолликул превращается в довольно крупный граафов
пузырек, внутри которого помещается яйцеклетка. В развитии фолликула
различают четыре фазы: созревание, овуляцию, образование на месте
овулировавшего фолликула желтого тела и фазу рассасывания желтого тела. От
количества овулировавших фолликулов и оплодотворенных в один период размножения
яйцеклеток зависит количество приплода.
В организме самки млекопитающих и в ее половом аппарате в
определенные периоды создаются благоприятные условия для оплодотворения
яйцеклеток и развития зародышей. По частоте возникновения в течение года этих
циклов млекопитающих разделяют на моноэстральных и полиэстральных.
Большинство животных, живущих на воле, размножается один раз в год и имеет один
период половой активности, во время которого животные спариваются. Вторую
группу составляют многие хозяйственно—полезные животные, большинство грызунов и
человек. У них в течение года циклы повторяются многократно.
Каждый половой цикл состоит из следующих фаз, или
стадий: предовуляционной, овуляционной, послеовуляционной, покоя. Каждая стадия
полового цикла характеризуется рядом изменений не только в половом аппарате, но
и в организме в целом.
У женщин половой цикл принято называть менструальным
циклом. Условно он определяется от первого дня наступившей менструации до
первого дня следующей менструации. Менструация возникает в послеовуляционном
периоде, если почему—либо не произошло оплодотворения, и на месте лопнувшего
фолликула вслед за овуляцией образуется эндокринная железа — желтое тело, продуцирующее
гормон прогестерон. С исчезновением желтого тела слизистая оболочка матки
подвергается обратному развитию. У высших обезьян и человека ее целостность
нарушается, в результате чего возникает кровотечение — менструация. Нормальной
длительностью менструального цикла у женщин считается 28 сут, продолжительность
менструаций — 2—3 сут.
При спаривании, или половом акте, в
половые пути самки вводится сперма самца. Половой акт представляет
сложнорефлекторный процесс, в нем участвует ряд безусловных и условных
рефлексов. У самцов и самок основными безусловными рефлексами являются: рефлекс
полового влечения (рефлекс отыскивания самки или самца), обнимательный рефлекс,
рефлекс подготовки половых органов (эрекция), совокупительный рефлекс, рефлекс
выделения гамет. В осуществлении безусловных половых рефлексов принимает
участие соматическая, симпатическая, парасимпатическая и отчасти
метасимпатическая нервная система.
У млекопитающих осеменение внутреннее, оно может быть
влагалищным и маточным. При первом эякулят попадает во влагалище, откуда
сперматозоиды, активно двигаясь, переходят в матку. При втором типе сперма
вводится непосредственно в матку. У человека влагалищный тип эякуляции.
Период, в течение которого сперматозоиды в половых путях самки сохраняют
способность к оплодотворению, как правило, незначителен. У человека он
ограничивается 2 сут. Оплодотворение происходит в основном в ближайшей к
яичнику трети яйцевода. Специфической особенностью оплодотворения является
проникновение сперматозоидов в цитоплазму яйцеклетки, слияние их ядер и
образование зиготы — оплодотворенной яйцеклетки, способной расти,
развиваться и давать начало новому организму.
Состояние самки от оплодотворения до рождения плода называют
беременностью. В эмбриональном развитии организма млекопитающих
различают зародышевый, предплодный и плодный периоды. У отдельных видов
животных развитие зародыша имеет ряд особенностей, касающихся характера
прикрепления эмбриона к стенке матки и условий его питания. Вначале зародыш
получает пищевые вещества с помощью осмоса. Этими веществами являются выделения
маточных желез, транссудат крови и лимфы. Позже питание происходит за счет
крови материнского организма. Последнее осуществляется через плаценту —
образование, развивающееся из наружной оболочки плода и слизистой оболочки
матки. Кровеносные сосуды плода и кровеносные сосуды матери входят в плаценте в
тесное соприкосновение, отделяясь друг от друга эндотелием капилляров. Кровь
матери и кровь плода, однако, никогда не смешиваются.
Плацента — не простая перегородка, она обладает сложными
биологическими функциями. Переход веществ из крови матери в кровь плода
происходит строго избирательно. Избирательная способность плаценты пропускать
одни вещества и задерживать или подвергать биологической обработке другие
получила название плацентарного барьера. Таким образом, на протяжении
всей беременности функционирует система мать—плацента—плод. Материнский
организм обеспечивает оптимальные условия для развития плода,
У плода процессы ассимиляции преобладают над процессами
диссимиляции, что обеспечивает его быстрый рост. Легкие у плода не функционируют,
из—за чего его кровообращение существенно отличается от кровообращения взрослых
организмов. Оно связано с плацентой и называется плацентарным. Кишечник у плода
начинает функционировать лишь во второй половине эмбрионального периода.
Движения плода прослеживаются в середине беременности. Головной мозг плода
находится в состоянии покоя, условнорефлекторной деятельности у него нет,
эндокринные железы функционируют.
Результатом законченного эмбрионального развития плода
являются роды. Они осуществляются благодаря сокращениям мускулатуры
матки и брюшного пресса. Процесс родов делят на три фазы: раскрытие родовых
путей, изгнание плода, послеродовая фаза. Родовой акт состоит из ряда
рефлексов, многие из которых начинаются с интероцепторов матки. В возникновении
родов наибольшая роль принадлежит нейрогуморальной и гормональной системе
матери и плацентарному комплексу. Во время третьей фазы родов выходят плодные
оболочки — послед.
Важнейшим анатомическим признаком млекопитающих животных и
человека являются молочные железы. Они относятся к системе органов
размножения. Образование молока после родов и его выведение из, молочных желез —
лактация — одно из важнейших биологических приспособлений. Молоко — естественная
пища новорожденных, которую, как указывал И. П. Павлов, приготовила сама
природа. Молоко имеет сложный химический состав и по биологической ценности
превосходит все другие продукты питания. В нем содержится более 100 различных
веществ, в том числе более 30 жирных кислот 20 аминокислот, 17 витаминов, много
ферментов, около 40 минеральных веществ и другие. Ряд соединений, такие как
казеин и лактоза, ни в каких других природных продуктах не встречаются.
Алексеева Л. В. Полицикличность размножения у приматов и антропогенез. М.,
1977.
Бабичев В. Н. Нейрогуморальная регуляция овариального цикла. М., 1984.
Биология развития млекопитающих. Методы / Под ред. М. Манка. М., 1990.
Вундер П. А. Эндокринология пола. М., 1980.
Герасимова Л. И., Лупандин Ю. В. Физиология репродукции. Петрозаводск,
1997.
Гормональная регуляция размножения у
млекопитающих / Под ред. К. Остина., Р. Шорта. М. 1987.
Грачев
И. И., Алексеев Н. П.
Роль рецепторов в регуляции лактации. Л., 1980.
Донован
Б. Т., Верф Тен Бош Дж. Физиология полового развития. М., 1974.
Држевецкая
И. А. Основы
физиологии обмена веществ и эндокринной системы. М., 1994.
Зеленская
Т. М. Эндокринные
взаимоотношения и тестикулярные антитела. Киев, 1981.
Новиков
С. Н. Феромоны и
размножение млекопитающих. Физиологические аспекты. Л. 1988.
Ноздрачев
А. Д., Чернышева М. П. Висцеральные рефлексы. Л., 1989.
Савченко
О. Н., Арутюнян Н. А., Степанов М. Г. Экспериментальное бесплодие: Эндокринологические аспекты.
СПб., 1992.
Савченков Ю. И.,
Лобынцев К. С.
Очерки физиологии и морфологии функциональной системы мать—плод. М., 1980.
Самойлов
А. П. Так
называемый секс. Л., 1991.
Смит
Дж. М. Эволюция
полового размножения. М., 1981.
Чернышева
М. П. Гормоны
животных. Введение в физиологическую эндокринологию. СПб. 1995.