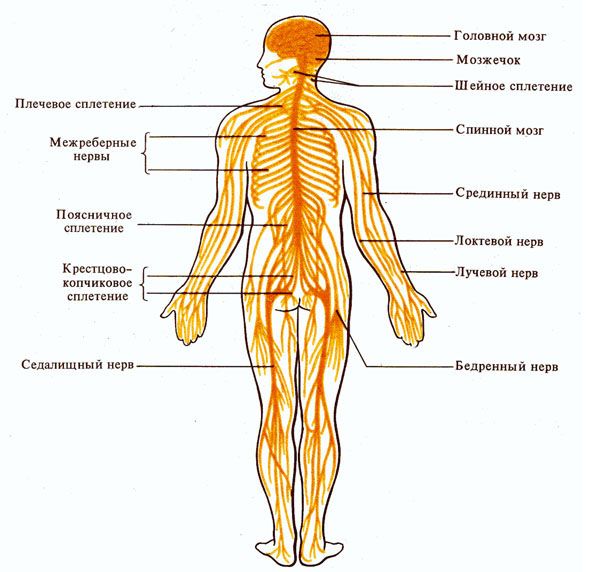
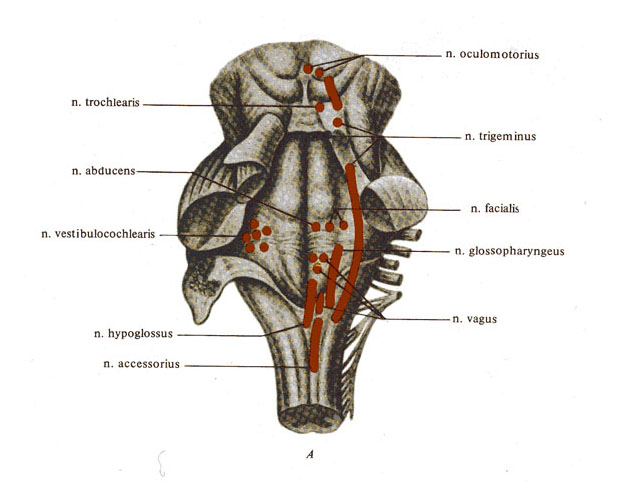
Рис. 264.
Центральная и периферическая нервная система.
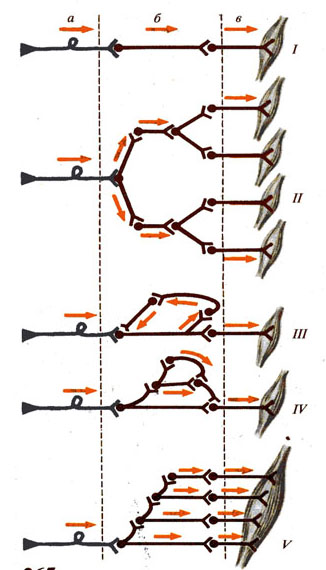
Рис. 265. Типы
нейронных связей в ЦНС (спинальные рефлекторные дуги):
I - линейная схема рефлекторной дуги, II - мультипликация нейронов, III - нейронная ловушка (субстрат памяти), IV - соотношение вставочных нейронов, обусловливающее трансформацию импульсов, V - вовлечение вышележащих нейронов и сегментов спинного мозга в распространение импульсов (иррадиация);
а - чувствительные нейроны, б - вставочные нейроны, в – мотонейроны.
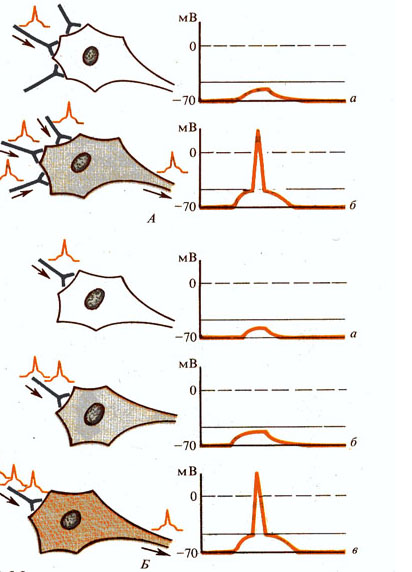
Рис. 266. Явление суммации:
А — пространственная суммация в результате одновременно наносимых раздражении: а — передача возбуждения с одного аксона (уменьшение мембранного потенциала), б — передача возбуждения с трех аксонов и генерация потенциала действия;
Б — временная суммация в результате последовательно наносимых раздражении: а - одно раздражение, б - два раздражения, в - три раздражения и генерация потенциала действия.
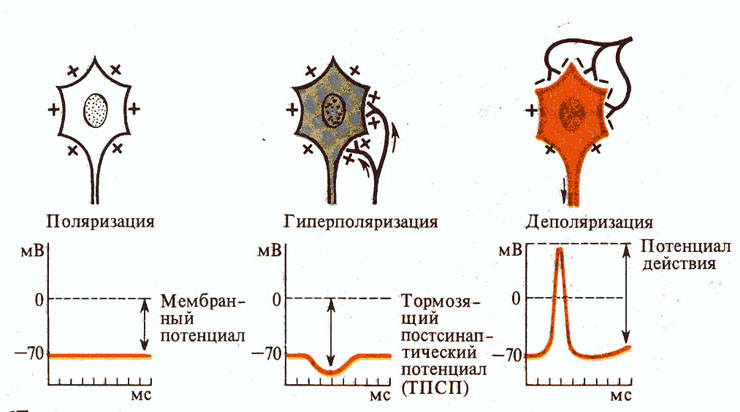
Рис. 267. Возможные состояния
нейрона.
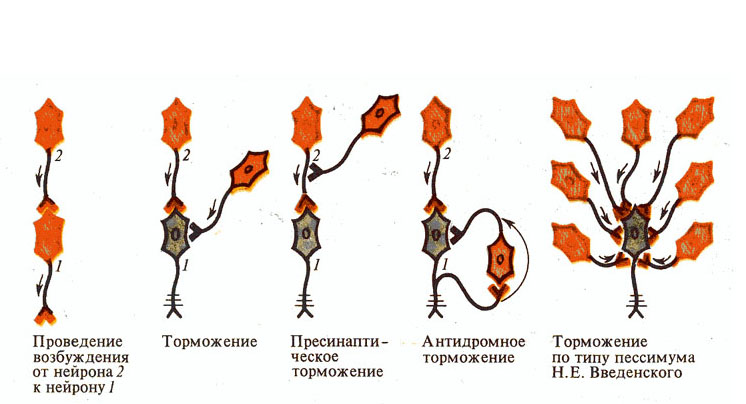
Рис. 268. Предполагаемые виды
торможения в ЦНС.
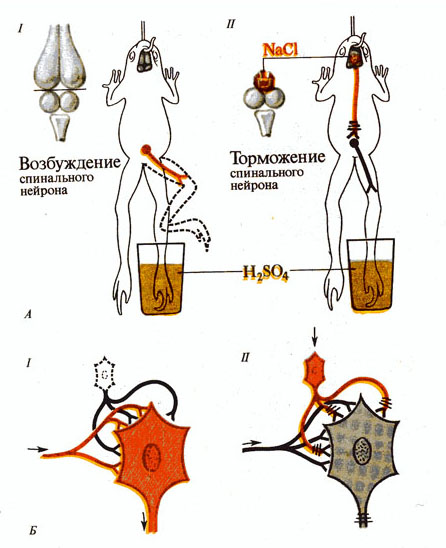
Рис. 269. «Сеченовское торможение»:
А - схема опыта: I - определение времени рефлекса у бесполушарной лягушки, II - увеличение времени рефлекса у той же лягушки после наложения кристаллика NaCI на область зрительных долей;
Б - предполагаемый механизм торможения: I - проведение возбуждения но мотонейрону, II - нисходящее тормозное влияние на мотонейрон (возбуждающие синапсы обозначены красным, тормозящие — черным).
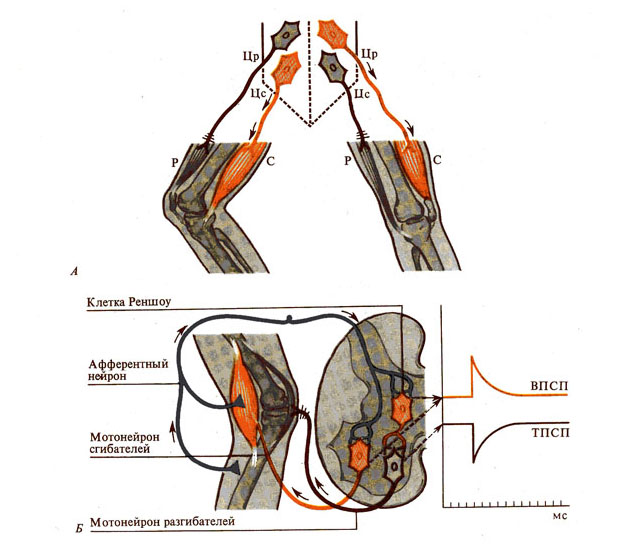
Рис. 270. Реципрокное торможение (по
Шеррингтону, 1897). А - состояние нижних конечностей (сгибание правой,
разгибание левой); Б - предполагаемый механизм торможения (передача возбуждения
на мотонейрон сгибателей и одновременное торможение мотонейpoнa
разгибателей
через клетки Реншоу):
Цр — центры разгибания в спинном мозге, Цс — центры сгибания, Р — мышцы-разгибатели, С — мышцы-сгибатели;
воэбужденные нейроны обозначены красным, тормозящие—серым.
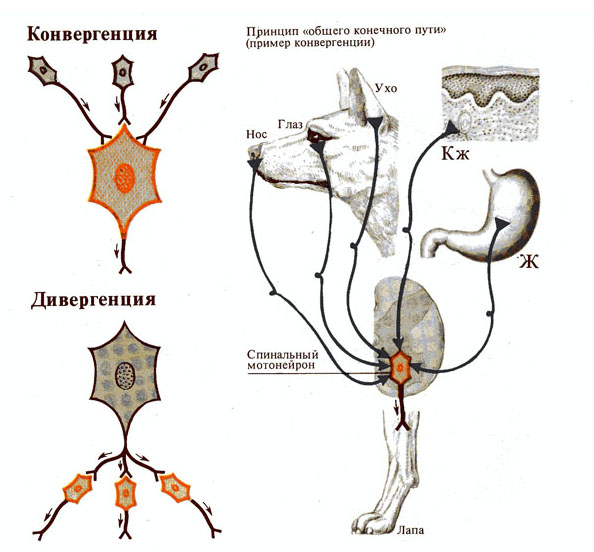
Рис. 271. Типы распространения
возбуждения в спинном мозге.
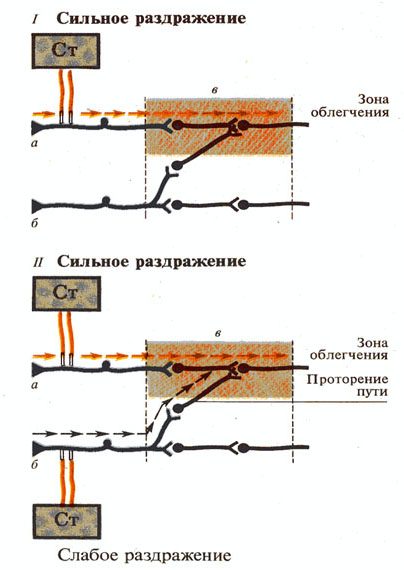
Рис. 272. Схема «проторения пути»: I - раздражение и возбуждение рефлекторной
дуги (а); б — невоэбужденная рефлекторная дуга; в — формирование зоны
облегчения; II -
присоединение раздражения рефлекторной дуги (б) и «проторение пути» в зону
облегчения.
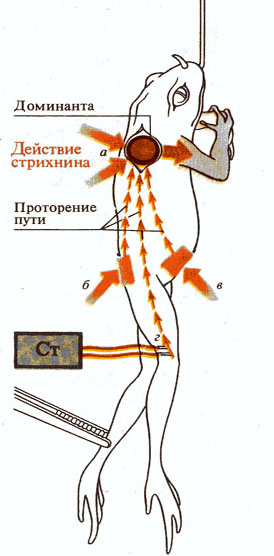
Рис. 273. Принцип доминанты (по А.
Ухтомскому 1923):
А - доминанта центров-сгибателей передних конечностей (при
аппликации стрихнина), б, в, г - рецептивные поля, раздражение которых
усиливает доминанту.
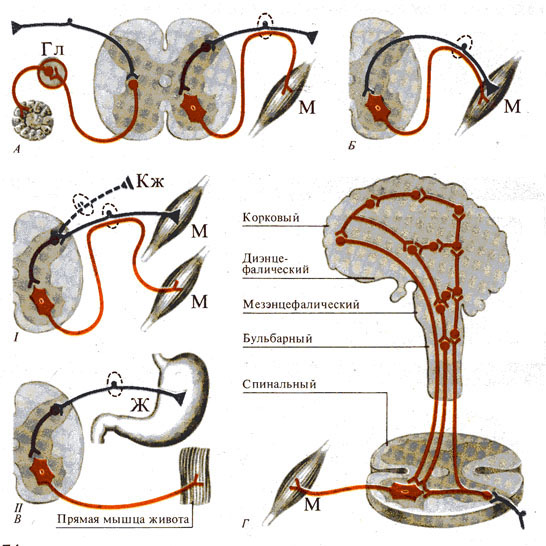
Рис. 274. Рефлекторная дуга. А - дуга
вегетативного (слева) и соматического (справа) рефлексов;
Б - гомонимная дуга; В - гетеронимная дуга с рецепторами,
заложенными в коже и мышцах ( I
)
и внутренних органах ( II ); Г- уровни «многоэтажной» рефлекторной дуги (по Э.
Асратяну, 1956)
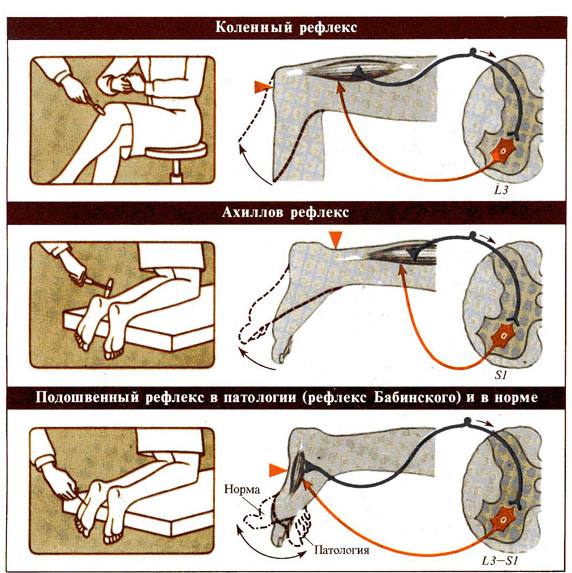
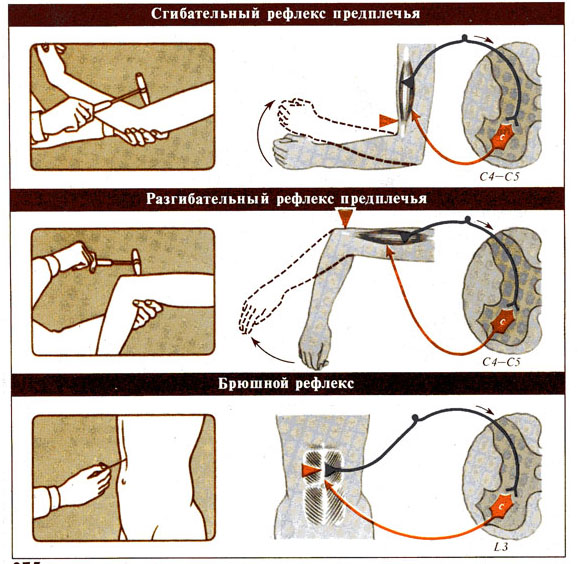
Рис. 275 А / 275
Б. Клинически важные рефлексы человека и локализация их центров в спинном
мозге.
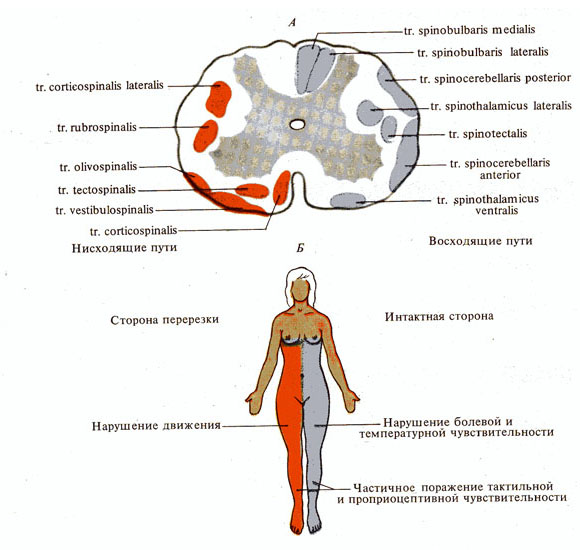
Рис. 276. Проводящие пути спинного
мозга. А - поперечный разрез спинного мозга; Б - зоны поражения у человека при
одностороннем повреждении спинного мозга на уровне грудных сегментов (паралич
Броун-Секара)
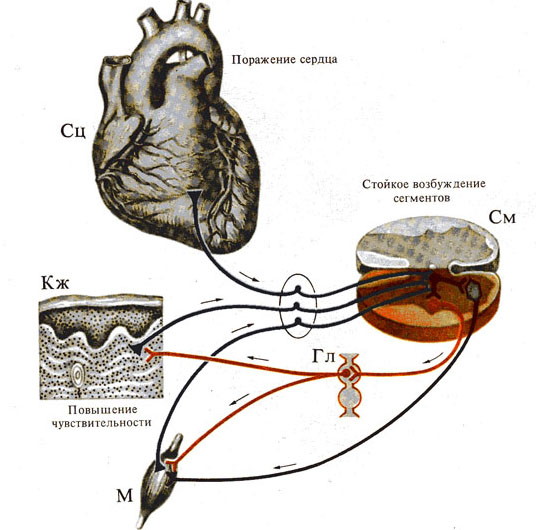
Рис. 277. Интероцептивные висцеро -
кутанный и висцеро - мускулярный рефлексы (отраженные боли в коже и мышцах при
патологии сердца)
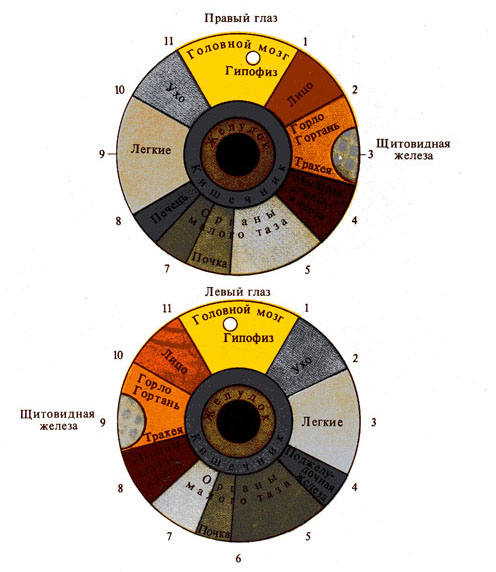
Рис. 278. Гипотетические проекционные
зоны тела человека на радужной оболочке глаза
(по Е. Вельховеру и др., 1981)
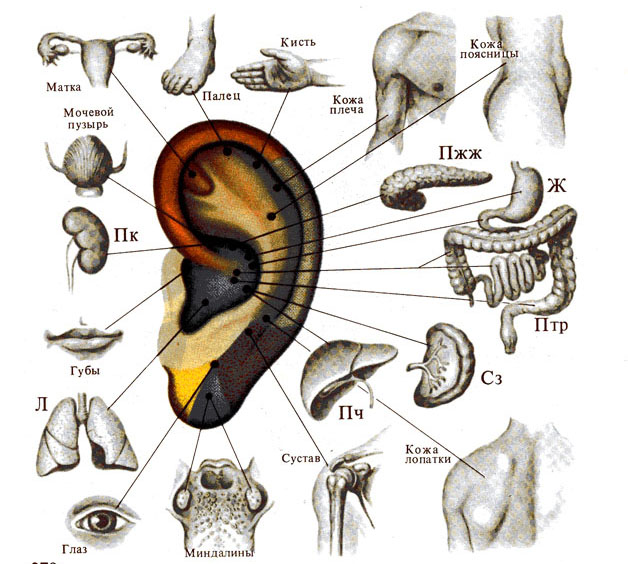
Рис. 279. Гипотетические проекции
некоторых внутренних органов и частей тела на ушную раковину (по В. Карягину,
1981).
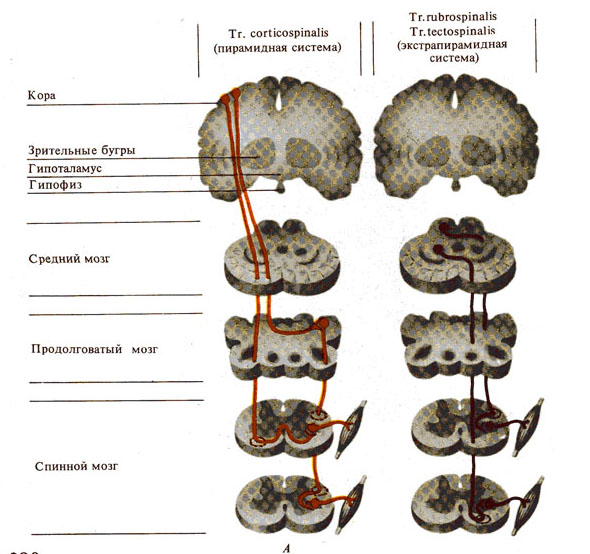
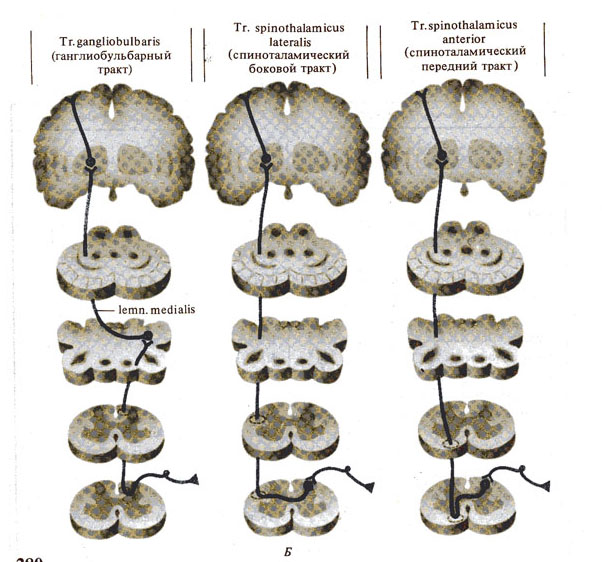
Рис. 280. Важнейшие проводящие пути спинного мозга. А - нисходящие, Б - восходящие.
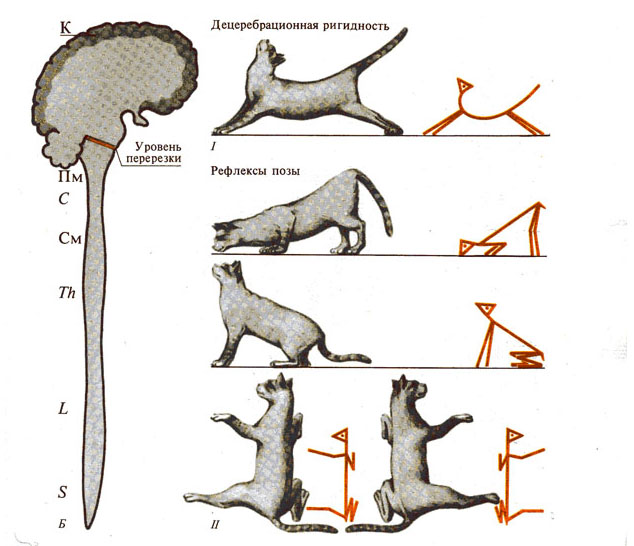
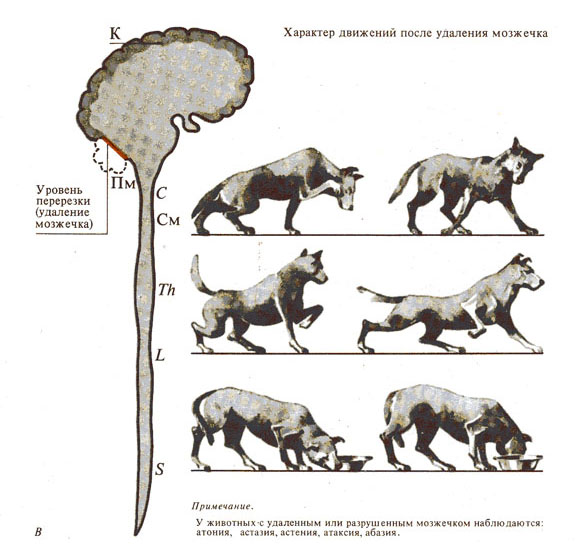
Таблица 1. Сравнительная характеристика тонической и моторной функций животных (кошки) с последовательным выключением различных отделов ЦНС.
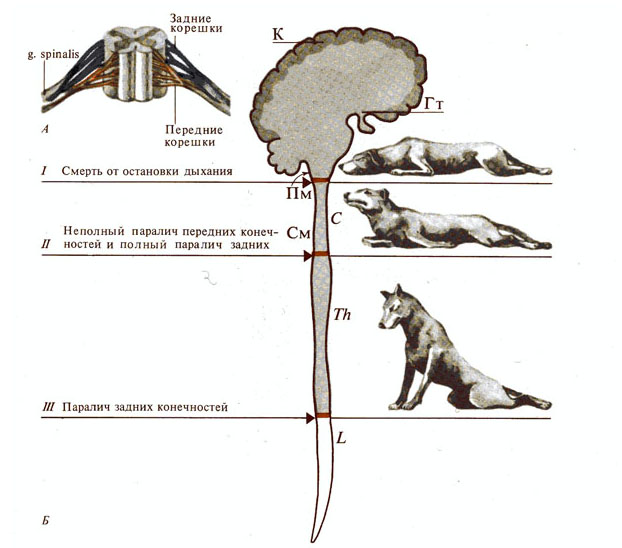
Рис. 281. Роль спинного мозга в
двигательных функциях. А - спинной мозг; Б - двигательные функции спинального
животного: I, II, III
-
уровни перерезок.
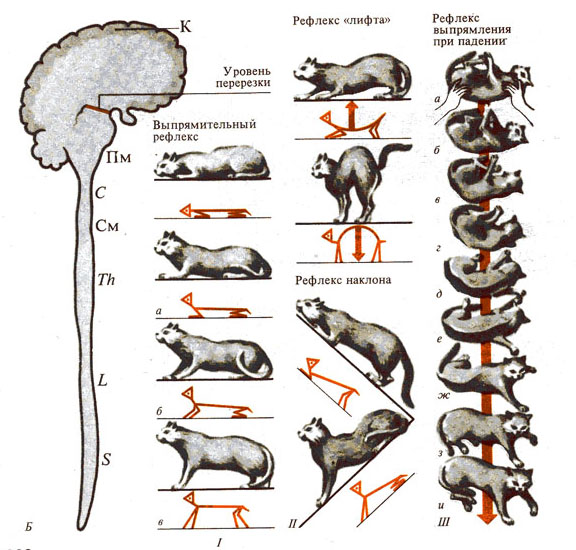
Рис. 282. Роль продолговатого мозга в двигательных функциях.
А - продолговатый мозг (разрез на уровне
четверохолмия); Б—двигательные функции бульбарного
животного: I - поза животного после перерезки ствола мозга ниже
уровня красных ядер, II - пассивные
статические рефлексы (зависимость тонуса сгибателей и разгибателей конечностей
от положения головы).
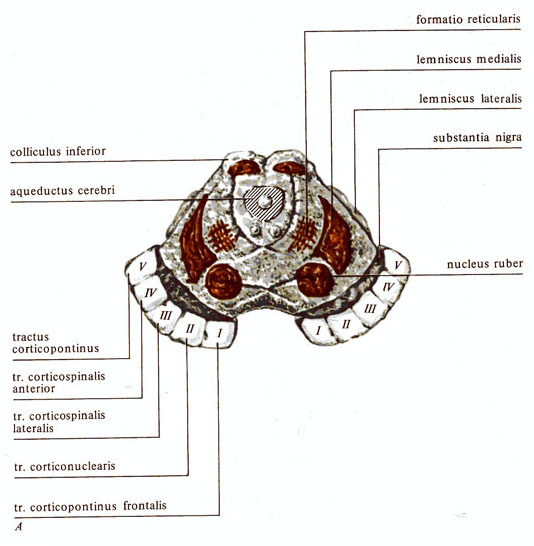
Рис. 283. Роль среднего мозга в двигательных функциях. А - средний мозг (разрез на уровне четверохолмия); Б - двигательные функции мезенцефалического животного: I - переход в стоячее
положение, II и IIIстатокинетические
рефлексы; а - и - этапы рефлекса.
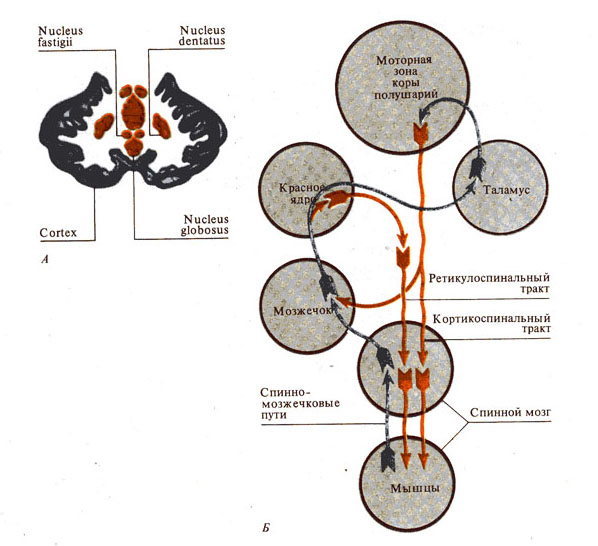
Рис. 284. Роль мозжечка в двигательных функциях.
А - основные структуры мозжечка
(разрез);
Б - схема связей мозжечка с другими
структурами мозга, обеспечивающими двигательные функции;
В - двигательные функции
безмозжечкового животного;
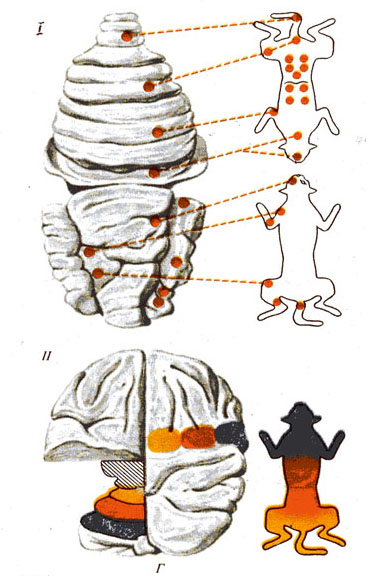
Г - локализация функций в мозжечке
(по А. Эдриану, 1943):
I – представительство различных рецепторных систем в мозжечке кошки; II - представительство функций в коре мозжечка: справа — области раздражения коры больших полушарий, слева - их проекции в мозжечке.
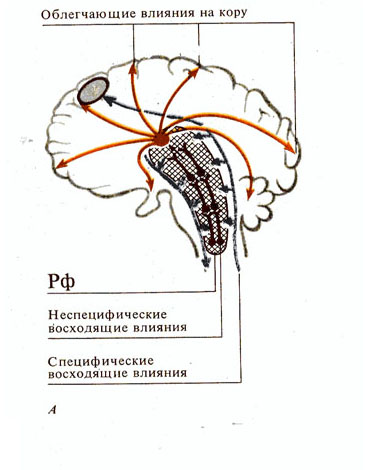
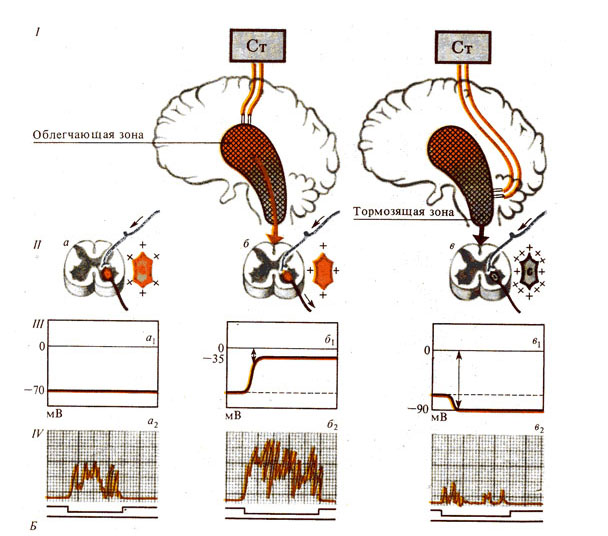
Рис. 285. Роль ретикулярной формации (Рф). А - восходящие влияния Рф; Б -
нисходящие влияния Рф и механизмы их реализации:
I - схема опыта раздражения облегчающей и тормозящей зон Рф; II - состояние мотонейронов спинного мозга: а - без раздражения Рф, б - при раздражении облегчающей зоны, в - при раздражении тормозящей зоны; III - состояние мембранного потенциала соответственно ( а1 – в1 ); IV - уровни ответной рефлекторной реакции, зависящей от состояния мотонейронов ( а2 – в2 )
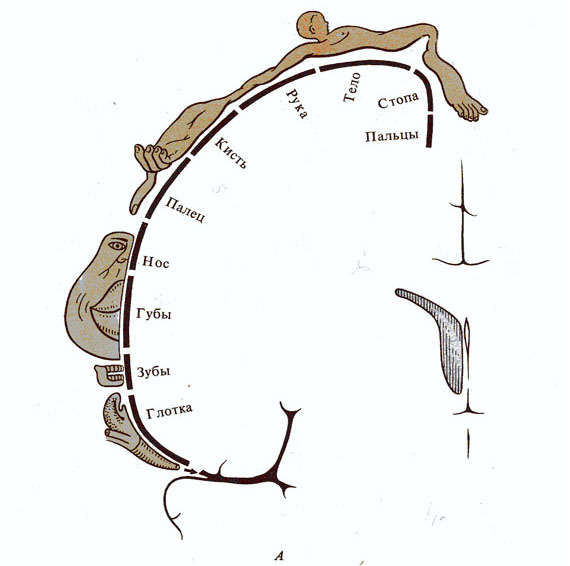
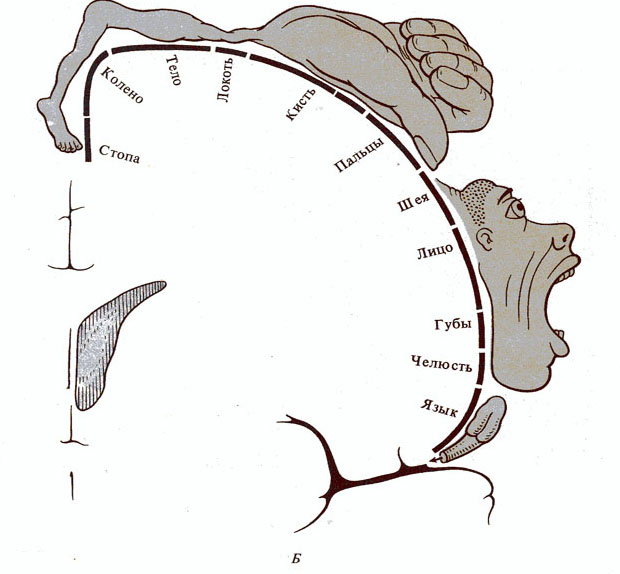
Рис. 286. Представительство чувствительных функций в задней
центральной извилине (А) и двигательных функций в
передней центральной извилине (Б). Части тела
гомункулюса соответствуют локализации данных функций в коре (по У. Пенфилду,
1956)
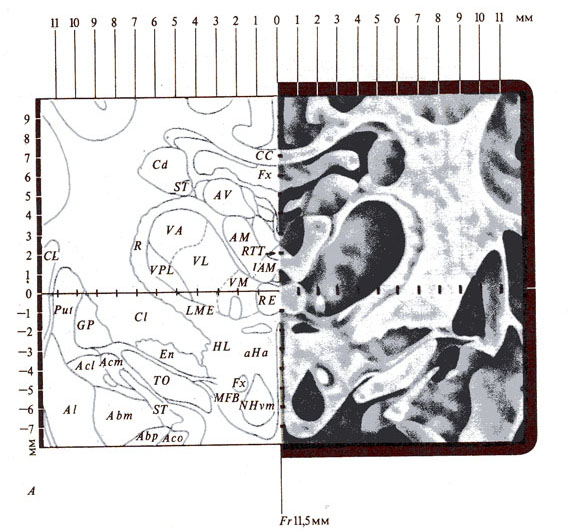
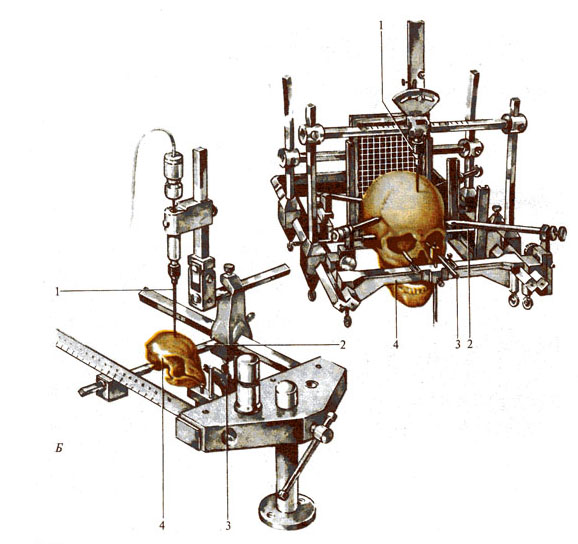
Рис. 287. Стереотаксическая техника как метод исследования
функций ЦНС. А - фронтальный срез мозга кошки с осями
координат и буквенными обозначениями подкорковых структур мозга (из атласа
Джаспера и Аймон-Марсана); Б - стереотаксические
установки для проведения опытов на животных и нейрохирургических операций на
мозге человека: 1 - электрод, 2 - ушные держатели, 3 - фиксаторы верхней
челюсти
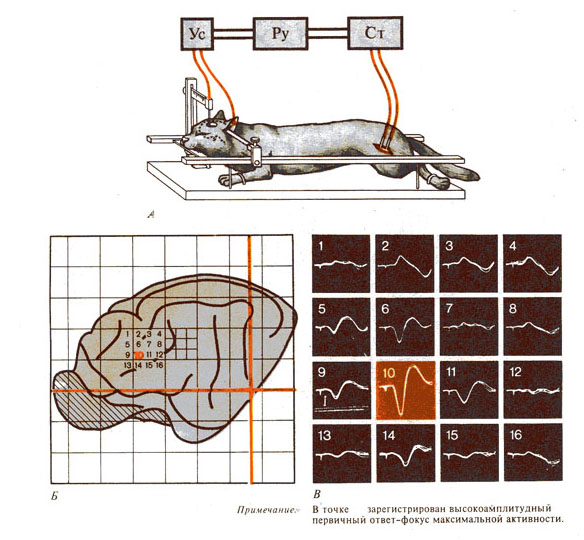
Рис. 288. Вызванные потенциалы (ВП) в
коре больших полушарий (по А. Башкирову, 1968). А - схема опыта; Б - карта коры
головного мозга кошки с точками регистрации ВП; В - ВП в точках регистрации.
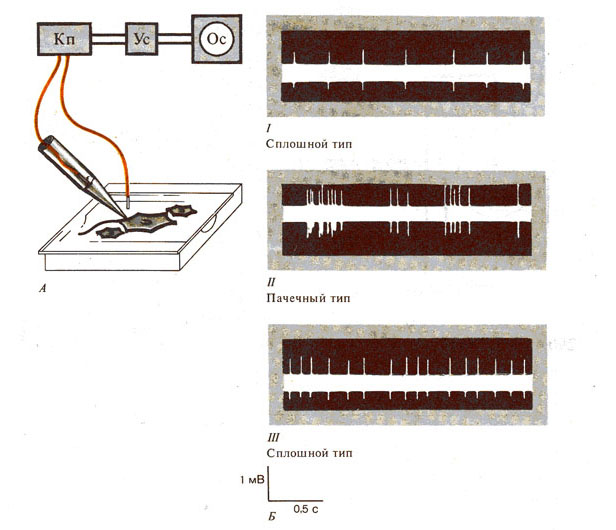
Рис. 289. Электрическая активность
отдельных нейронов и ее регистрация (внеклеточное отведение) (по И. Власовой,
1982). А — схема опыта; Б — типы спайковой активности нейронов различных
отделов мозга: I - активность нейронов зрительной коры, II - активность нейронов
гиппокампа, III - клеток Пуркине, Кп - катодный повторитель.
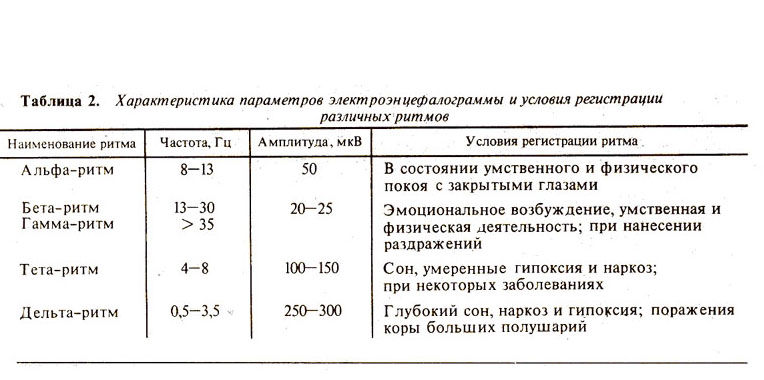
Таблица 2. Характеристика параметров
электроэнцефалограммы и условия регистрации различных ритмов.
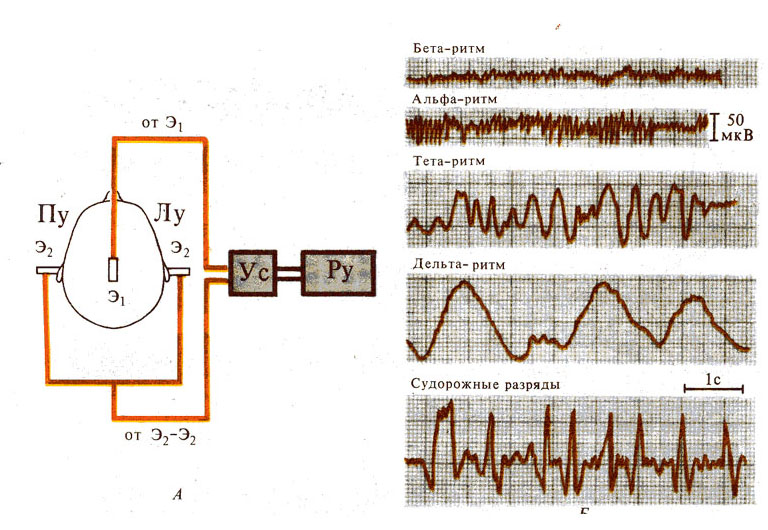
Рис. 290. Электроэнцефалография. А —
схема регистрации ЭЭГ; Б — основные ритмы ЭЭГ: Э1 - активный
электрод, Э2 - индифферентные электроды, ПУ и ЛУ — правое и левое
ухо
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}