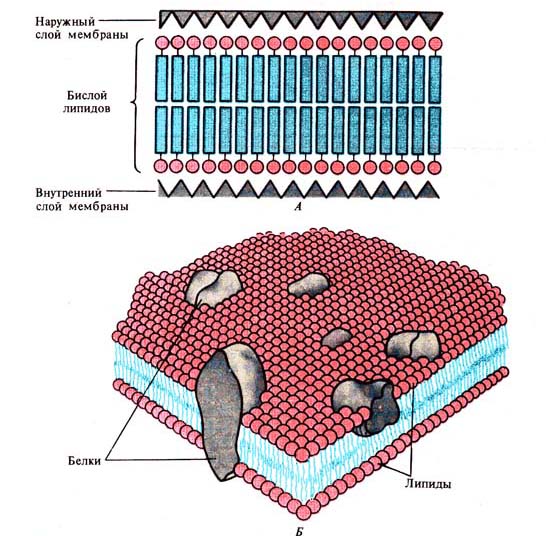
Рис. 233. Схема строения возбудимых
мембран. А — гипотетическая структура клеточной мембраны (по И. Даниели, 1950);
Б — мозаичная модель клеточной мембраны (по Л. Бергельсону, 1975)
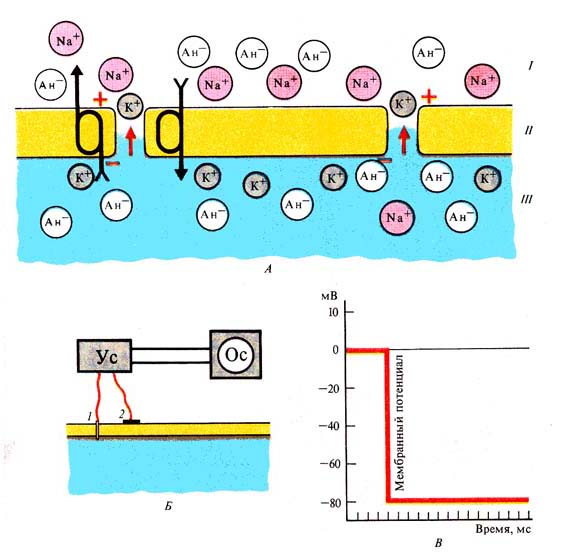
Рис. 234. Мембранный потенциал:
А — поляризация мембраны за счет деятельности Nа+,
К+ -помпы; / — внешняя среда, // — мембрана, III — содержимое
клетки; Б — схема регистрации мембранного потенциала: / — электрод внутри
волокна, 2 — поверхностный электрод; В — графическое изображение мембранного
потенциала
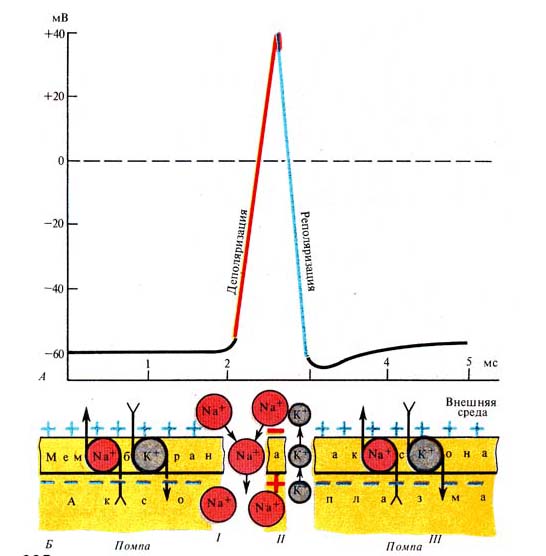
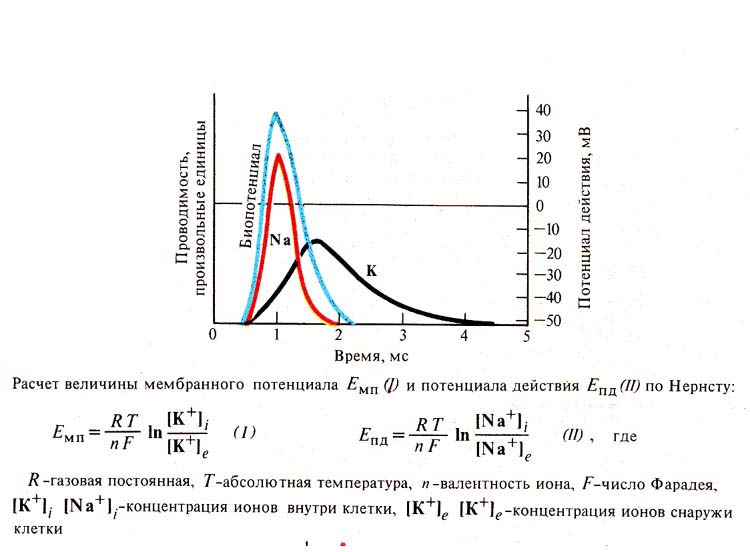
Рис. 235. Сопоставление развития
потенциала действия (А) с изменениями проницаемости мембраны (Б) (по К.
Кулланде, 1968): I — нарушение
деятельности Nа+, К+ -помпы, изменение проницаемости
мембраны, вхождение ионов Nа внутрь клетки и изменение заряда мембраны (деполяризация):
II - выход ионов К наружу
(реполяризация); III - возобновление деятельности Nа+, К+
-помпы
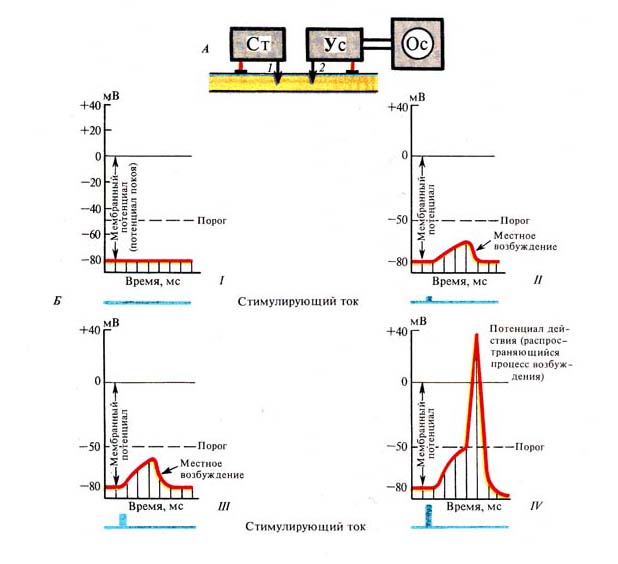
Рис. 236. Изменение мембранного
потенциала и формирование потенциала действия в зависимости от силы
раздражения: А — схема опыта: 1 — стимулирующий, 2 — отводящий микроэлектроды;
Б— графики регистрации потенциала действия: I—потенциал
покои до раздражения, //, ///—местное возбуждение (раздражение током
возрастающей силы), IV—потенциал действия—бегущая волна возбуждения
Рис. 237. Интенсивность диффузии ионов
Nа внутрь волокна и ионов К наружу в сопоставлении с биопотенциалом нервного
волокна
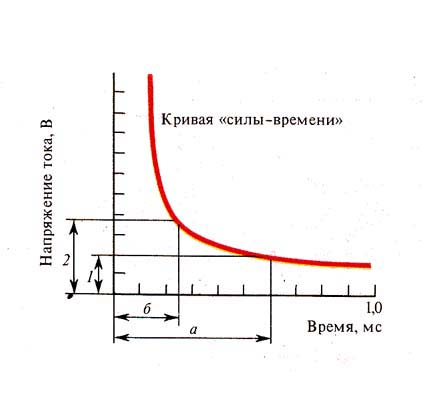
Рис. 238. Кривая «силы-времени» (по
Лапику и др., 1926): / — реобаза, 2 — удвоенная реобаза; а — полезное время
действия тока, б — хронаксия
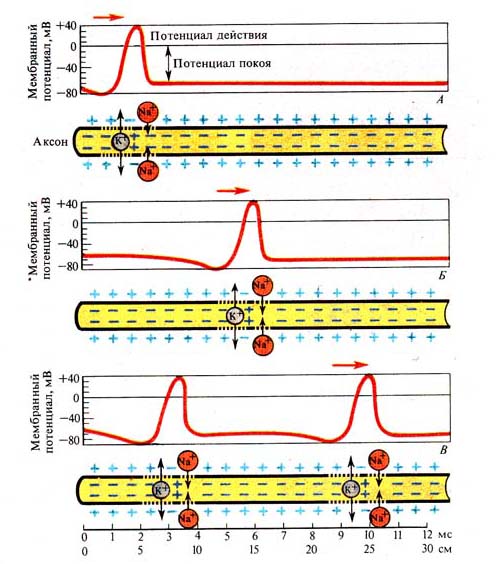
Рис. 239. Этапы распространения
возбуждения по аксону нервной клетки
(А, Б, В) (по Б. Катцу и Е. Жукову, 1970)
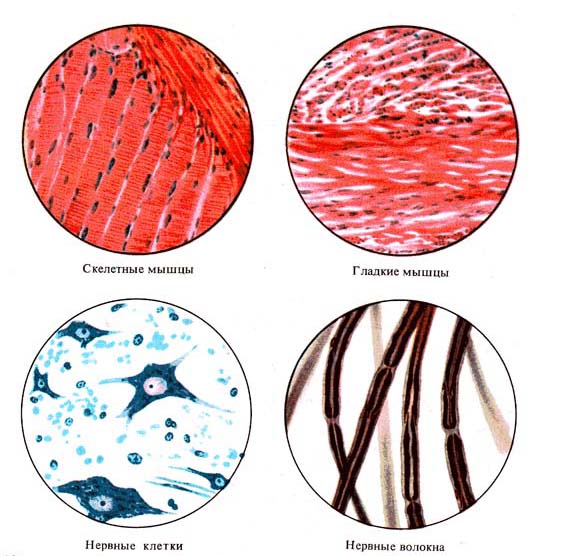
Рис. 240. Возбудимые ткани
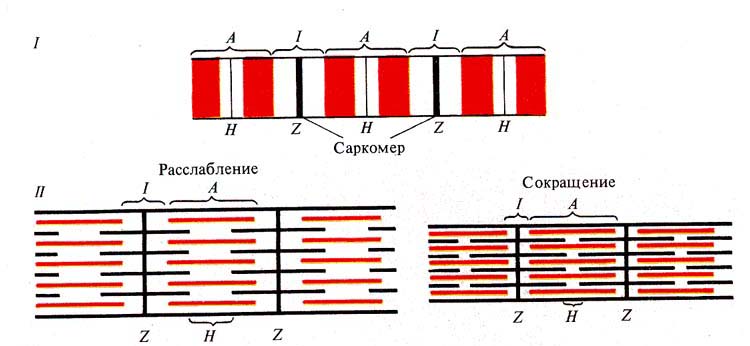
Рис. 241. Строение мышечного волокна
(I) и миофибриллы (II):
А — анизотропные диски, / — изотропные диски, Н и Z — пластинки
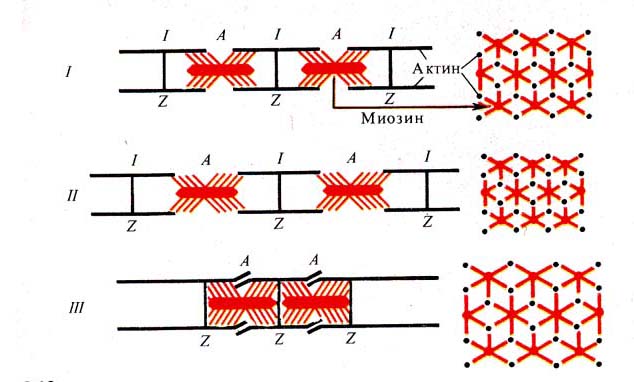
Рис. 242. Схема скольжения нитей
актина и миозина в процессе сокращения (по
Дж. Бендоллу, 1970):
/ — расслабленная миофибрилла, // — растянутая миофибрилла,
/// — сокращенная миофибрилла, слева — продольное расположение нитей, справа —
поперечное сечение
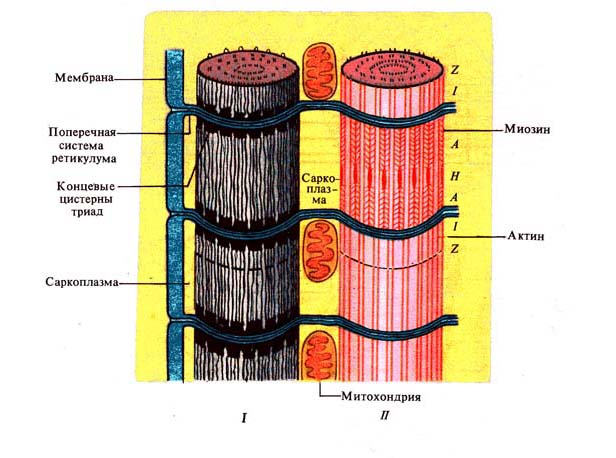
Рис. 243. Миофибриллы с
саркоплазматическим ретикулумом (по Дж.
Бендоллу, 1970):
/ — саркомеры, // — саркомеры с удаленным ретикулумом; А —
анизотропные диски, / — изотропные диски, Н и Z — пластинки
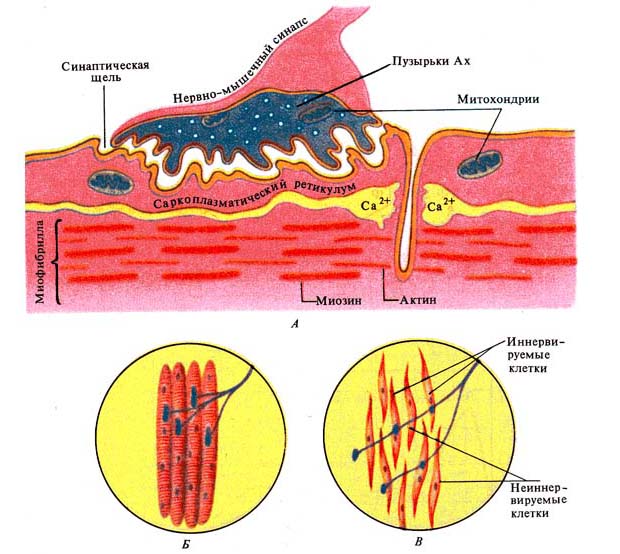
Рис. 244. Строение нервно-мышечного
синапса. А -— нервно-мышечный синапс в разрезе;
Б — распределение синапсов соматического аксона в волокнах
скелетной мышцы; В — распределение синапсов симпатического аксона в клетках
гладкой мышцы
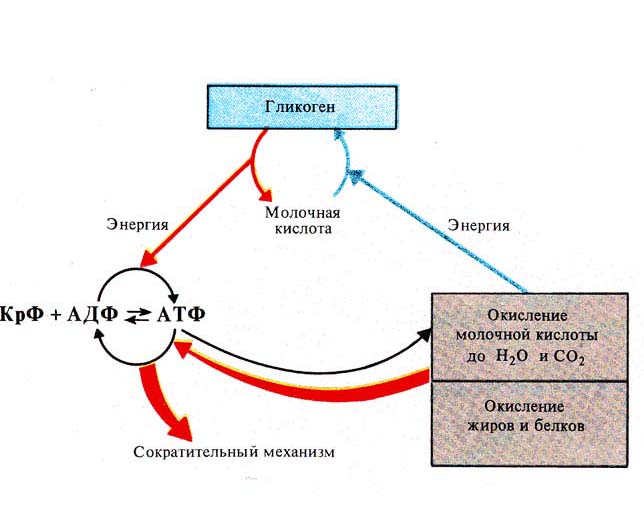
Рис. 245. Передача на рабочий механизм
мышечного волокна энергии, освобождающейся при окислительных процессах
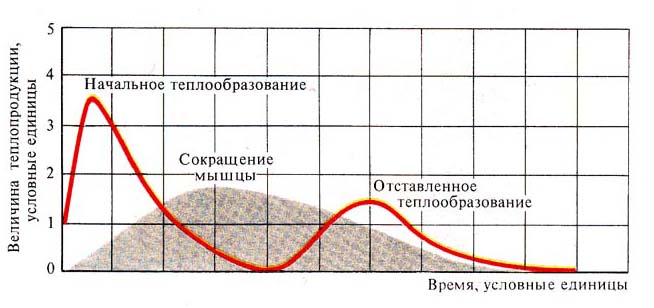
Рис. 246. График теплопродукции мышцы
в процессе одиночного сокращения
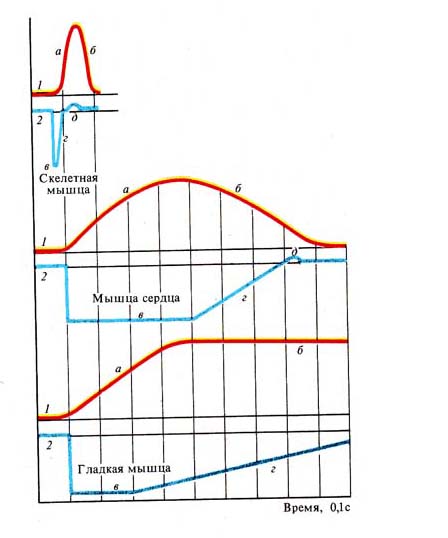
Рис. 247. Сократимость и возбудимость
мышц разного вида: / — кривая сокращения, 2 — кривая возбудимости; а — период
укорочения, б — период расслабления, абсолютный рефракторный период, и —
относительный рефракторный период, д — фаза экзальтации
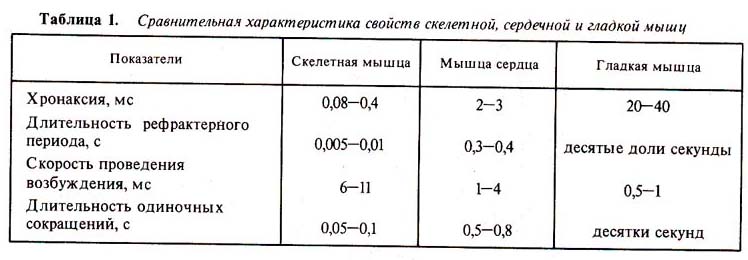
Таблица 1. Сравнительная характеристика свойств
скелетной, сердечной и гладкой мышц
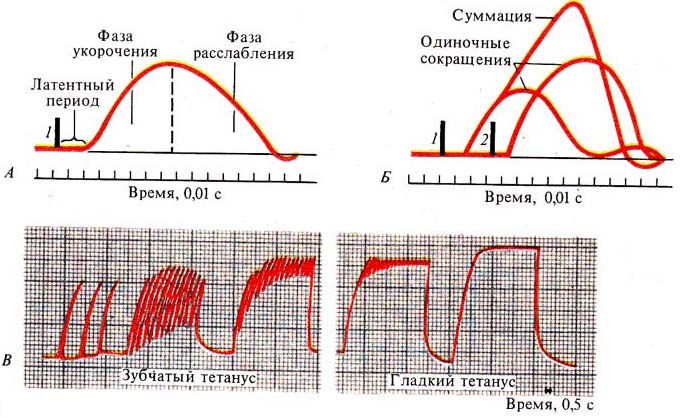
Рис. 248. Одиночное сокращение
(А), суммация (Б), тетанус (В): I — момент первого раздражения,
2 — момент второго раздражения
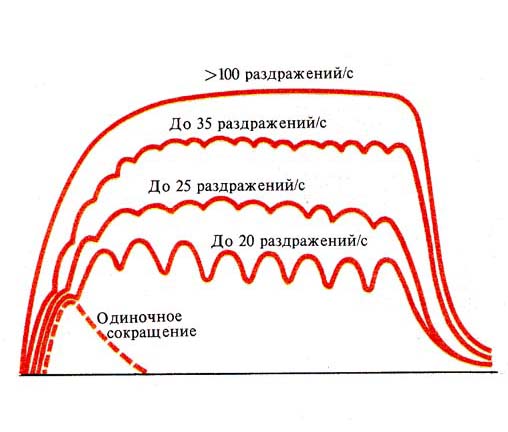
Рис. 249. Формирование тетануса в
зависимости от частоты раздражения
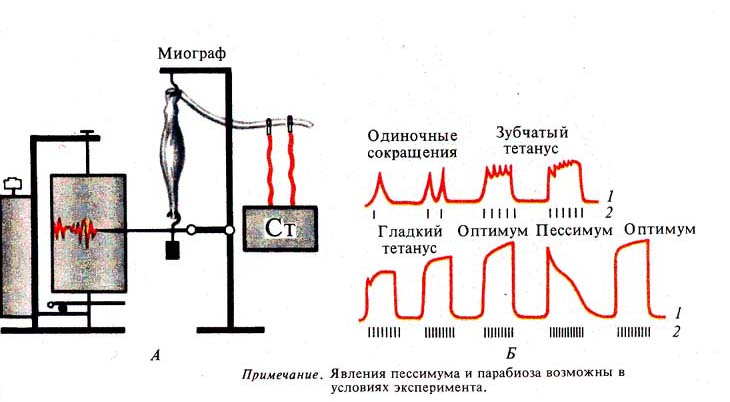
Рис. 250. Оптимум и пессимум (по Н.
Введенскому). А — схема регистрации; Б — кривые мышечных сокращений (1) при различной частоте раздражении (2)
Рис. 251. Парабиоз (по Н.
Введенскому):
А — схема опыта: / — положение электродов, // — создание
очага парабиоза;
Б —кривые мышечных сокращений (тетанусы) (1) при нарастающей
силе тока (2)
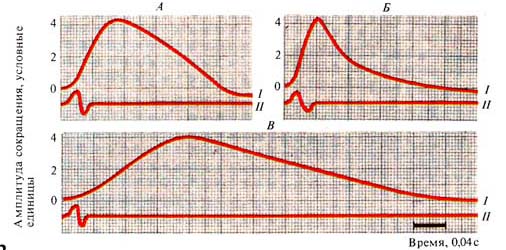
Рис. 252. Кривые одиночного сокращения
смешанной мышцы (А), мышцы из белых волокон (Б), мышцы из красных волокон (В);
I —
сокращение, // — ток действия
Рис. 253. Строение двигательной
единицы
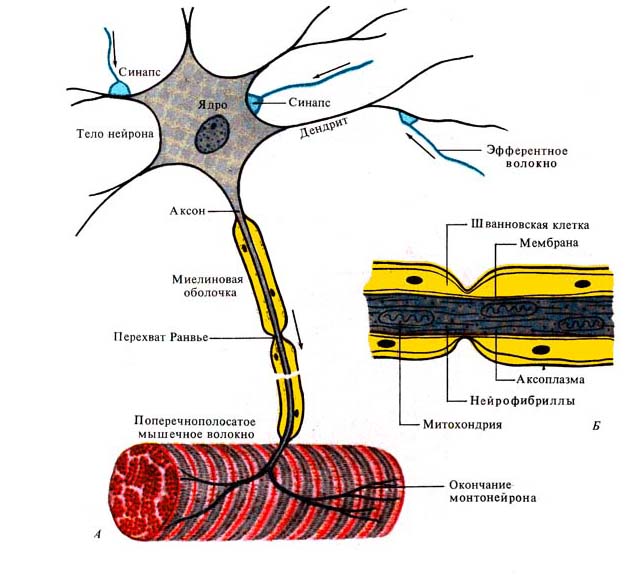
Рис. 254. Нейрон и его компоненты. А —
нервная клетка, аксон, мышца; Б — строение нервного волокна
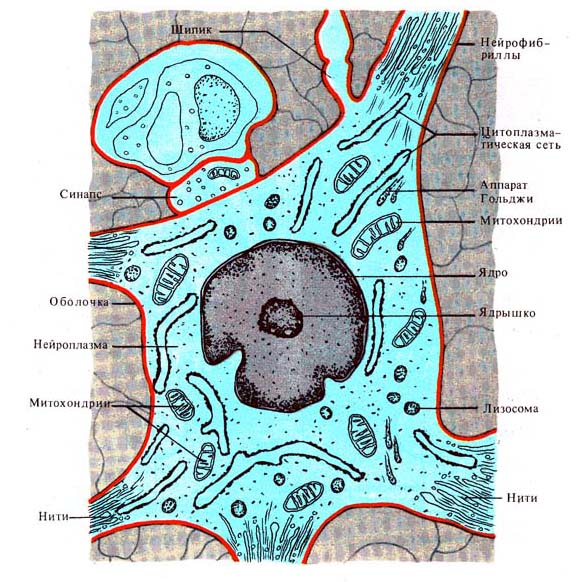
Рис. 255. Нейрон (схема электронной
микрофотографии) (по А. Миколадзе, 1970)
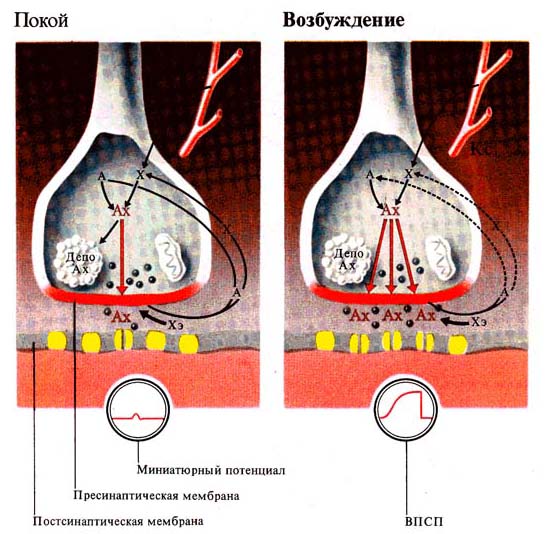
Рис. 256. Синаптические процессы в
невозбужденном и возбужденном синапсах (по Л. Щельцыну, 1980):
А — ацетат, Х — холин, Хэ — холинэстераза
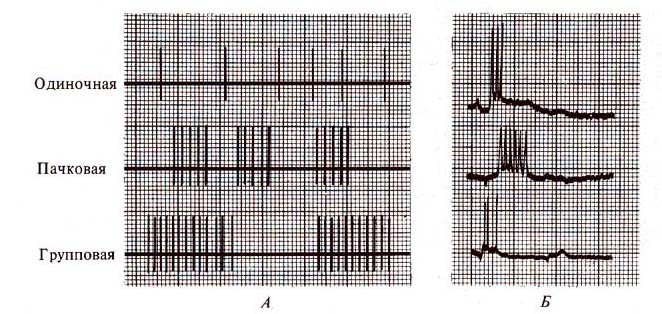
Рис. 257. Фоновая и вызванная
импульсная активность нейронов (по Л. Шельцыну, 1980). А — типы активности
ретикулярного нейрона; Б — типы вызванных ответов нейронов при внутриклеточной
регистрации
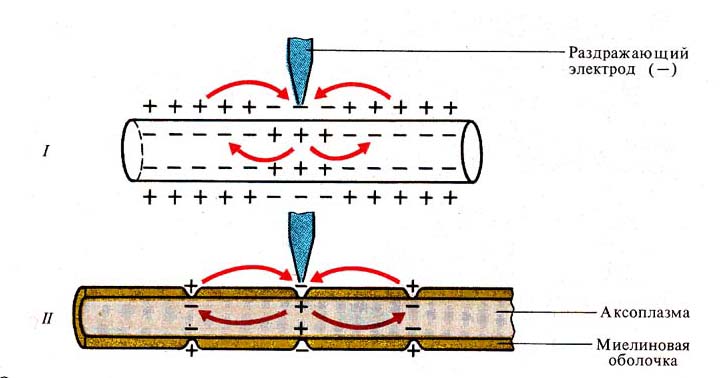
Рис. 258. Проведение возбуждения в
нервных волокнах (по Дж. Бендоллу, 1970):
I —
немиелинизированное волокно
// — миелинизированное
волокно (сальтаторное проведение)
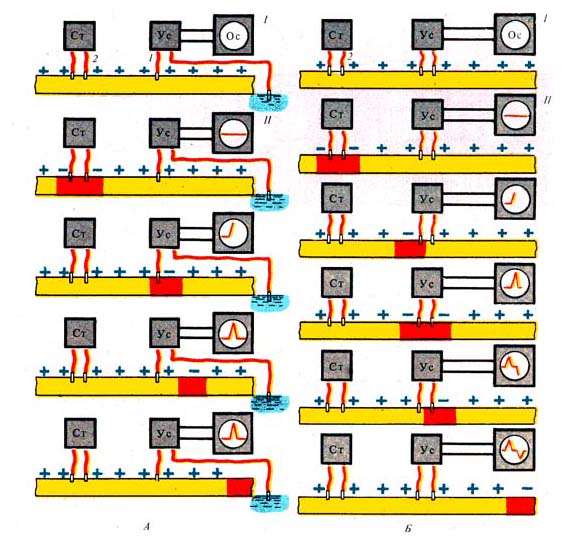
Рис. 259. Регистрация потенциала
действия нервного волокна (по К. Кулланде, 1968). А — униполярное отведение
(однофазный потенциал); Б — биполярное отведение (двухфазный потенциал):
/ — схема опыта, // — схема формирования потенциалов; 1 —
отводящие электроды, 2 — раздражающие электроды
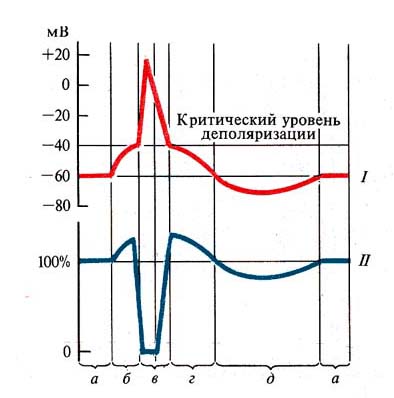
Рис. 260. Сопоставление одиночного
возбуждения (/) с фазами возбудимости (//):
а — мембранный потенциал (исходная возбудимость), б —
локальный ответ, или ВПСП (повышенная возбудимость), в — потенциал действия
(абсолютная и относительная рефрактерность), г — следовая деполяризация
(супернормальная возбудимость), д — следовая гиперполяризация (субнормальная
возбудимость)
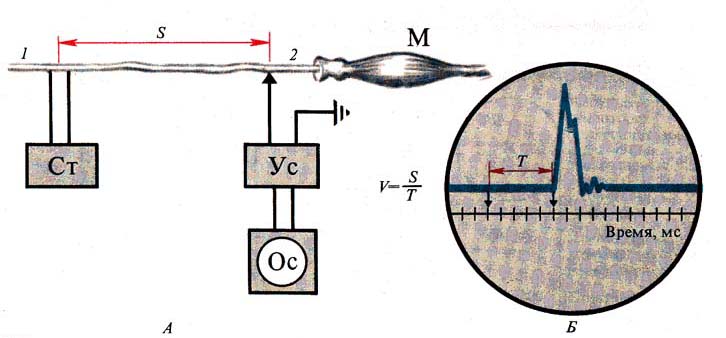
Рис. 261. Измерение скорости
проведения возбуждения по нервным волокнам. А — схема опыта; Б — осциллограмма:
V — скорость проведения возбуждения, S — расстояние между раздражающими (1) и отводящими
(2) электродами, Т — время между моментом раздражения и моментом прихода волны
возбуждения к отводящему электроду (2)
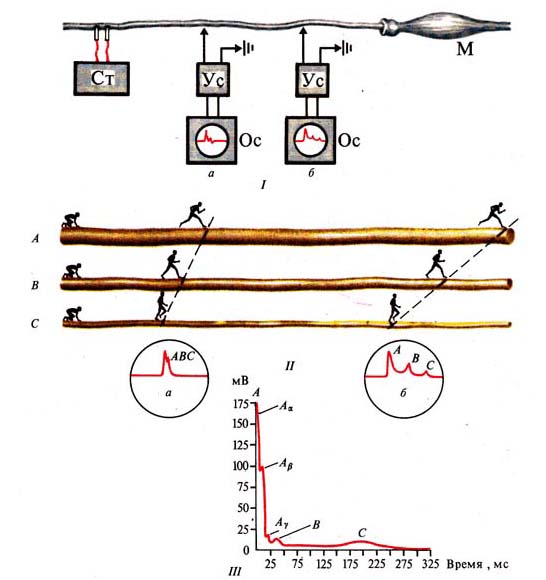
Рис. 262. Скорость распространения
возбуждения по нервным волокнам разного типа: /—//—схема Опыта: а — установка,
регистрирующая потенциалы нерва на небольшом
расстоянии от раздражающих электродов, б — установка, регистрирующая
потенциал нерва на большом расстоянии от раздражающих электродов (человечками
обозначены импульсы);
/// — соотношение компонентов потенциала действия нерва,
содержащего А-, В-, С-типы нервных волокон (по Гассеру и Эрлангеру, 1937)
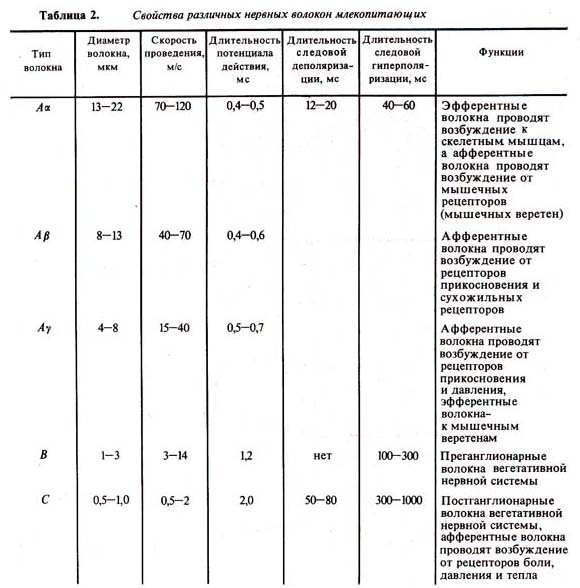
Таблица 2. Свойства различных нервных волокон млекопитающих
Рис. 263. Физиологический электротон.
А — схема опыта; Б — изменение мембранного потенциала (а), критического уровня
деполяризации (б) и возбудимости (в) при действии на возбудимую ткань
постоянного тока (стрелкой отмечено начало действия тока)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}